Research progress on skin photoaging mechanisms and natural extracts
Jieyong Lai
1,2,3,#

,
Xingxuan Ren
1,2,3,#
,
Gaobin Liang
4
,
Shuhui Jia
1,2,3
,
Jiapeng Li
1,2
,
Yuwei Wang
1,2
,
Huang Zhang
1,2
,
Weidong Xie
1,2,3,*
*Correspondence to:
Weidong Xie, State Key Laboratory of Chemical Oncogenomics, Shenzhen International Graduate School, Tsinghua University, Shenzhen 518055, Guangdong, China.
E-mail: xiewd@sz.tsinghua.edu.cn
Ageing Cancer Res Treat. 2026;3:202605. 10.70401/acrt.2026.0027
Received: January 23, 2026Accepted: June 16, 2026Published: June 17, 2026
Abstract
Skin photoaging is a progressive, ultraviolet (UV)-driven form of extrinsic skin aging that compromises epidermal barrier integrity, dermal extracellular matrix organization, pigmentary homeostasis, and subcutaneous tissue support. Current findings indicate that photoaging is driven by interconnected molecular mechanisms, including ultraviolet A-(UVA)- and ultraviolet B (UVB)-induced reactive oxygen species (ROS) generation, DNA photolesions, mitogen-activated protein kinase/activator protein-1 (MAPK/AP-1-) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)-mediated inflammatory signaling, matrix metalloproteinase activation, collagen and elastin degradation, mitochondrial dysfunction, autophagy impairment, and senescence-associated secretory phenotypes (SASP). Plant-derived polyphenols, carotenoids, and terpenoids primarily attenuate oxidative stress and inflammatory signaling; marine-derived mycosporine-like amino acids, sulfated polysaccharides, xanthophylls, and collagen peptides provide UV absorption, matrix protection, and structural support; and microbiome-derived metabolites and probiotics modulate redox balance, immune signaling, and barrier homeostasis via the gut-skin axis. By integrating layer-specific pathogenesis with multi-source natural interventions, this review highlights translationally relevant strategies for developing safer, mechanism-guided anti-photoaging therapies, informing both preclinical research and clinical applications.
Keywords
Skin photoaging, ultraviolet radiation, oxidative stress, inflammation, natural products
1. Introduction
The human skin and its appendages cover about 1.5-2.0 m2 of surface and are the first line of defense against external insults such as pathogens, chemicals, ultraviolet radiation (UVR) and physical injury. It also helps to control body temperature, conserve water and mediate sensory perception[1-4]. Skin aging arises from intrinsic and extrinsic processes. Intrinsic aging is driven by endogenous factors, including genetic predisposition and cellular metabolism, and manifests as reductions in epidermal thickness, collagen and elastin content, vascularity, and cell proliferation, collectively diminishing skin elasticity and mechanical resilience[5-7].
Extrinsic aging, commonly referred to as photoaging, results primarily from chronic exposure to ultraviolet radiation, with clinical features including fine and coarse wrinkles, dyspigmentation, rough texture, laxity, telangiectasia, and impaired barrier recovery[6,8,9]. At the molecular level, photoaging involves oxidative stress, inflammatory signaling, extracellular matrix (ECM) degradation, and dysregulation of dermal fibroblast function, largely mediated by matrix metalloproteinases (MMPs) and pro-inflammatory proteases[10].
The biological consequences of ultraviolet (UV) exposure are wavelength- and depth-dependent[10,11]. Ultraviolet B (UVB) radiation (280-315 nm) is absorbed mainly in the epidermis and directly induces cyclobutane pyrimidine dimers and 6-4 photoproducts in keratinocytes and melanocytes[12-14]. Ultraviolet A (UVA) radiation (315-400 nm) penetrates more deeply into the dermis, where it promotes oxidative stress, mitochondrial injury, vascular and immune alterations, and extracellular matrix remodeling[11,15]. Recent evidence indicates that high-energy visible light (blue light) also contributes to photoaging by inducing reactive oxygen species (ROS) and Opsin-3-mediated melanogenesis, leading to persistent hyperpigmentation, particularly in darker skin phototypes[16].
At the molecular level, these events converge on several mutually reinforcing pathways. ROS generation activates mitogen-activated protein kinase/activator protein-1 (MAPK/AP-1) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling, increases cytokine and matrix metalloproteinase expression, suppresses collagen synthesis, and accelerates fibroblast senescence. Persistent DNA damage, mitochondrial dysfunction, autophagy dysregulation, and senescence-associated secretory phenotypes (SASP) further amplify chronic inflammation and tissue degeneration[17-19]. Although many studies have described individual mechanisms of photoaging, fewer reviews integrate tissue-layer specificity with molecular signaling and natural extract-based interventions. This integration is important because epidermal DNA damage and dermal matrix breakdown are biologically distinct but clinically interconnected components of photoaged skin[10,19,20].
Natural bioactive materials have therefore attracted increasing interest as complementary or preventive interventions[21,22]. Plants, marine, and microbial extracts have become potential options, exhibiting the ability to maintain dermal architecture with the help of strong antioxidant and anti-inflammatory properties[23-25].
A structured literature search was conducted in PubMed, Web of Science, Scopus, Embase, and Google Scholar for articles published between January 2000 and May 2026. Search terms included “skin photoaging”, “ultraviolet radiation”, “UVA”, “UVB”, “reactive oxygen species”, “matrix metalloproteinases”, “extracellular matrix”, “mitochondrial dysfunction”, “autophagy”, “natural products”, “plant extracts”, “marine-derived compounds”, and “skin microbiome”. Primary research, mechanistic, translational, and clinical studies were prioritized, with reviews used for contextual background. This review synthesizes tissue-layer specific pathogenesis with natural extract-based interventions, providing a mechanism-oriented perspective for photoprotection in dermatology and cosmetic science.
2. Skin Structure and Layer-Specific Responses to Photoaging
2.1 Skin anatomy
Skin is a multilayered organ consisting of the epidermis, dermis, and subcutaneous tissue, with each layer providing particular protection against mechanical forces, chemical attacks, microbial attacks, and ultraviolet radiation[26-28]. The epidermis is a stratified squamous epithelium with keratinocytes, and their spatial organization defines successive steps of differentiation, starting with the basal layer up to the spinous, granular and the cornified strata. This vertical organization points to a slow and tightly regulated differentiation process that often receives limited attention until it is disrupted under pathological conditions. Among the keratinocyte population are intercalated melanocytes that regulate pigmentation, Langerhans cells that carry out immune surveillance, and Merkel cells that are involved in signaling by mechanoreceptors[14,26]. The structural proteins (filaggrin and filaggrin-2) together with tight junctions and intercellular lipids are the main structural proteins in the barriers of this organizational tier. Together, these parts inhibit the transepithelial water loss and block the xenobiotics and UVB radiation ingress, even though the barrier was imperfect[29-32].
The dermis is a connective tissue under the epidermis, which is functionally integrated but structurally heterogeneous, and which is more active than a mere supportive scaffold. It has scattered fibroblasts that are embedded in a collagen and elastin-rich mesh which gives the skin tensile strength and elasticity, and is involved in the metabolic maintenance of the epidermis[33-35]. In addition to anchorage, the dermis has also been shown to be a signaling interface via a dermal epidermal junction and has a certain level of control over epidermal turnover as well as plays a role in the long-term maintenance of barrier homeostasis[14,33].
The subcutaneous tissue forms the deepest layer and consists predominantly of adipocytes interwoven with connective tissue. While less visible, its functional contribution is substantial, extending from mechanical cushioning to thermoregulation[36-38]. When the effects of ultraviolet exposure are considered across these layers, the resulting damage is unevenly distributed. UVB radiation primarily affects superficial compartments, where it disrupts epidermal DNA and compromises barrier integrity. In contrast, UVA penetrates more deeply, influencing the dermal matrix as well as immune and vascular components[27,39,40]. Photoaging therefore does not arise from isolated lesions alone but reflects layer-specific patterns of damage that develop within the structural organization of the skin[27,39,40].
2.2 The epidermal layer
The epidermis is the outermost and most directly UV-exposed compartment of the skin. Most UVB photons are absorbed in this layer and generate DNA photolesions, including cyclobutane pyrimidine dimers (CPDs) and 6-4 photoproducts, particularly in basal keratinocytes and melanocytes[27,41]. These lesions activate canonical genome-protective pathways, including the p53/p21 and p16INK4a/Rb signaling axes, leading to transient cell-cycle arrest, DNA repair, or apoptosis. Persistent exposure, however, promotes keratinocyte senescence and reduces epidermal regenerative capacity[41,42].
Besides direct genotoxicity, UV irradiation induces oxidative stress in keratinocytes, thus activating MAPK, NF-κB, and nuclear factor erythroid 2-related factor 2 (Nrf2) pathways, which cascade to cause inflammatory responses, antioxidant defenses, and differentiation programs[43-45]. Long-term exposure to UVB leads to impaired epidermal barrier integrity, which is manifested by increased transepidermal water loss (TEWL) and dysregulation of filaggrin, tight-junction protein, and cornified envelope protein expression[26,30,42]. Melanocytes dynamically increase the melanin synthesis to cover nuclear DNA; however, photo-flattened melanocytes accumulate in the cutaneous tissues of the senescent type, which leads to the formation of heterogeneous pigmentation and mottled dyschromia[14,26]. At the same time, the occurrence of UVB-induced cytokine and chemokine secretion by keratinocytes reduces the Langerhans cell density and activity, creating the localized immunosuppression and low-grade chronic inflammation[14,46]. Besides, blue-light exposure may trigger Opsin-3-dependent melanogenesis, augmenting pigmentation in epidermal cells[16]. All these perturbations together undermine the integrity of the epidermis and provide a substrate that sup-ports the following destruction of the dermis[42,47].
2.3 The dermal layer
The dermis is the principal structural compartment affected by UVA-induced photoaging. Because of its longer wavelength, UVA penetrates into the dermis and generates reactive oxygen species that damage nuclear DNA, mitochondrial DNA, proteins, and lipids[26,27,48]. The resulting oxidative burden activates activator protein 1 (AP-1) and NF-κB, increases matrix metalloproteinase (MMP)-1, MMP-3, and MMP-9 expression, and disrupts collagen and elastin organization[26,48,49].
UVA has been shown to induce mitochondrial dysfunction specifically in the dermal fibroblast, that leads to reduced collagen synthesis and SASP[48,50,51]. In a paracrine manner, senescent fibroblasts secrete pro-inflammatory cytokines, growth factors, and proteases, causing chronic inflammation and ECM degradation to become self-enhancing[19,20,52]. Recent 2026 studies further highlight that UVA-driven mitochondrial stress exacerbates mitochondrial DNA (mtDNA) instability and ROS accumulation, reinforcing SASP-mediated dermal inflammation and matrix remodeling[53,54]. Histologically, the photoaged dermis is characterized by isolated collagen bundles, accumulation of abnormal elastic fibers, higher vascularity density, and immune cells infiltration[27,35,55]. Such accumulated changes compromise the mechanical strength of the skin, which is clinically evident in deep wrinkles, laxity, and sagging[56-58].
2.4 The subcutaneous tissue
Subcutaneous adipose tissue not only offers mechanical and metabolic support to the dermis, but also its importance is often underestimated[59,60]. This loss of fat that is caused by senescence and UV irradiation leads to loss of skin thickness and the appearance of wrinkles[37,61]. It has been shown empirically that UV irradiation suppresses lipid production by inhibiting the adipogenic transcription factors, and the inhibition is achieved following a single exposure event[37,61].
UV irradiation also exerts marked modulatory effects on adipokine secretion from adipocytes: it induces a massive reduction in adiponectin secretion, along with a massive increase in adipokine expression, particularly the pro-inflammatory cytokine interleukin-6 (IL-6). The imbalance between adipokines and cytokines disrupts the physiological function of dermal fibroblasts besides creating immense microenvironmental conditions in the skin, leading to cutaneous chronic pro-inflammatory settings[37,59,62,63].
At the same time, age-related changes in the cellular structure of the immune compartment in the subcutaneous stratum further hinder the process of adipocyte renewal, effectively slowing down the process of hypodermal atrophy[37,64]. These metabolic abnormalities and structural impairment, when intertwined together, weaken the biomechanical structures of the skin, eventually causing skin turgor loss, the final outcome of photoaged skin.
2.5 UVA and UVB skin layer response
The biological effects of UVA and UVB differ with distinct photochemical actions and maximal depths of penetration. This localized damage is clinically characterized by acute erythema and, on a molecular scale, by the coordination of the DNA repair programs and pro-inflammatory cascades, thus outlining an unmistakably mechanistic divergence from the indirect, oxidative-stress-induced effects of UVA[18,65,66]. In models, UVB induces epidermal hyperplasia and increases wrinkles and is precipitated by a secondary loss of dermal collagen[59]. Similarly, other studies support these observations and report parallel hyperplastic and wrinkle-inducing effects of UVB irradiation[39].
On the contrary, UVA enters the deep dermis and subcutaneous tissue, where it mainly triggers oxidative stress, but does not necessarily produce DNA photolesions[36,67,68]. This oxidative attack leads to mitochondrial DNA damage, accumulation of the common deletion and long-term activation of the MMPs to cause sustained dermal remodeling and skin laxity in the presence of no acute inflammation[18,69]. It therefore follows that although the two wavelengths play a role in photoaging, UVB plays a central role in epidermal pathology and inflammatory consequences, whereas UVA is the major cause of chronic dermal loss and biomechanical senescence[36,67,70].
3. Molecular Mechanisms of Skin Photoaging
Since chronic exposure to UV light initiates a cascade of biochemical and cellular changes that interact and occur in a coordinated and cumulative manner, the molecular pathogenesis of skin photoaging includes UV-induced oxidative stress, ECM degradation, chronic inflammation, genomic instability, mitochondrial impairment and dysregulated autophagy, thereby remodeling the structural and functional integrity of epidermal and dermal compartments[15,20,52]. Despite being mediated by distinct signaling pathways, these processes are closely interdependent, forming a self-perpetuating cascade that increasingly disrupts skin homeostasis[36,71].
3.1 Reactive oxygen species production and oxidative stress
UVR containing both UVA and UVB wavelengths induces skin photoaging mainly through the induction of ROS. These highly reactive molecules are formed when UV photons interact with endogenous chromophores and mitochondrial electron transport chains in the cutaneous cells[43,72,73]. The resultant photochemical excitation then causes the production of superoxide anions, hydroxyl radicals, and hydrogen peroxide and so overwhelms the intrinsic antioxidant defenses, most notably manganese superoxide dismutase (MnSOD). Consequently, the state of oxidative stress is established, which is characterized by an imbalance between the production and clearance of ROS[72-75].
The excess ROS cause oxidative damage to the cellular macromolecules, lipid, proteins, and nucleic acids, causing membrane integrity, enzyme activities, and genomic stability to be disrupted. The combined effects of these events compromise the homeostasis and viability of the cell[72,73,76]. Additionally, ROS are the key secondary messengers that stimulate redox-sensitive signaling pathways, including the phosphoinositide 3-kinase/protein kinase B (PI3K/Akt) and NF-κB cascades, which in turn feed inflammatory responses and can also regulate gene expression profiles related to photoaging[73,74]. The mitochondrial electron transport chain is both a source and a target of ROS, with the mutations of mitochondrial DNA triggered by UV and the loss of membrane potential accelerating the production of ROS in a feed-forward loop, which increases oxidative stress and enhances cellular senescence and apoptosis[15,75,77].
The UV-ROS-oxidative stress axis is therefore an essential molecular pathway that leads to the cumulative effects of photodamage and premature skin aging by triggering an intricate network of oxidative stress, inflammation signaling, and mitochondrial dysfunction[15,72,74].
3.2 Extracellular matrix degradation
The MAPK/AP-1 signaling is a major driver in relaying collagen and ECM degradation during skin photoaging through activation of transcriptional induction of MMPs, notably MMP-1, which degrades fibrillar collagens that are indispensable for dermal structural support[76,78,79]. UV-generated ROS activate MAPKs (e.g., p38, ERK, and JNK), that phosphorylate transcription factors like AP-1, which consists of c-Fos and c-Jun subunits; upon activation, the AP-1 complex translocates into the nucleus to interact with promoter sites found in MMP genes, promoting their expression[72,78,80]. The subsequent overexpression of MMPs results in the degradation of collagen types I and III and other ECM constituents, leading to dermal scaffold disruption and loss of skin elasticity/resilience, which is clinically expressed as wrinkle formation or skin firmness[72,81,82].
In human skin, experimental treatments such as administration of phytochemicals like syringaresinol have shown inhibition of MAPK phosphorylation and AP-1 activation, leading to decreases in MMP-1 production and maintenance of collagen synthesis, emphasizing the potential therapeutic use of targeting this pathway for treatment against photoaging[83,84]. Furthermore, the MAPK/AP-1 and other signaling pathways, such as NF-κB and transforming growth factor β (TGF-β)/Smad cascades, are also involved in inflammatory response and collagen biosynthesis, forming an intricate network for ECM remodeling upon UV-mediated harm[82,85,86]. This model illuminates the importance of MAPK/AP-1-mediated MMP stimulation as a central highway in the molecular pathogenesis of facial photoaging, and connects oxidative stress with architectural ECM damage via stringent transcriptional modulation[72,82,85].
3.3 Inflammatory signaling pathways
Inflammation plays an important role in skin photoaging and is closely associated with an increase in the SASP. Senescent cells acquire SASP, a versatile secretory program characterized by the release of pro-inflammatory cytokines, chemokines, growth factors, and metalloproteinases. Through these secreted molecules, SASP remodels the tissue microenvironment, promoting chronic inflammation, extracellular matrix remodeling, and paracrine signaling, which may exacerbate photoaging phenotypes and contribute to age-related tissue dysfunction[87].
NF-κB is a central inflammatory pathway through which UVR and oxidative stress induce the transcription of pro-inflammatory mediators, including IL-6, tumor necrosis factor-alpha (TNF-α), and cyclooxygenase-2 (COX-2)[74,88,89]. Activated by upstream signals such as toll-like receptor (TLR) engagement or oxidative stress, the inhibitor of κB (IκB) is phosphorylated and subsequently degraded by the proteasome. This process releases NF-κB dimers, primarily composed of p65/p50, which bind to κB sites on target gene promoters and initiate transcription of pro-inflammatory mediators[88,89].
As a result, IL-6 and TNF-α accumulate, acting as autocrine and paracrine effectors that amplify inflammatory cascades. In addition, COX-2, an inducible enzyme responsible for prostaglandin synthesis, further enhances inflammation and contributes to tissue remodeling[74,90,91]. NF-κB activation is tightly regulated and can interact with signaling crosstalk involving MAPKs and protein kinase C theta (PKCθ), which modulate the timing and magnitude of cytokine expression. This regulation is critical for NF-κB nuclear translocation and DNA binding specificity[92,93].
Pharmacologically, inhibition of IκB phosphorylation or upstream kinases suppresses NF-κB activation and reduces IL-6, TNF-α, and COX-2 expression. These findings underscore the central role of NF-κB in inflammatory pathophysiology and highlight it as a potential therapeutic target[74,94]. Overall, NF-κB-directed cytokine expression represents a tightly controlled molecular pathway that integrates cellular and tissue responses to extracellular stress, linking upstream signals to downstream gene-expression programs[74,88,89].
3.4 DNA damage and cellular senescence
The DNA damage response (DDR) in skin photoaging is strictly regulated by a complex network of tumor suppressor proteins and cell cycle checkpoint regulators, with p53, p16INK4a, p21Cip1/Waf1, and SASP playing predominant roles in influencing cellular outcomes upon exposure to genotoxic stress[15,95,96]. Thereafter, when UV irradiation damages DNA, p53 is rapidly stabilized and activated through a series of chemical modifications that enable it to function as a transcription factor[15,95]. The first step is to arrest the cell cycle progression at G1/S with the help of p21 activation, allowing sufficient time for DNA repair. This is followed by induced apoptosis of cells with extensive DNA damage to avoid mutations[97-99].
Simultaneously, the cyclin-dependent kinase (CDK) inhibitor p16 is an important regulator of the retinoblastoma (Rb) pathway, which enhances cell cycle arrest, and promotes the development of a stable senescence state that is marked by permanent growth arrest and insensitivity to mitogenic cues[100,101]. The interaction between p53-controlled cell-cycle checkpoints and p16-controlled senescence pathways provides a fail-safe mechanism ensuring that no DNA-damaged cells proliferate. p21 acts as a dynamic controller of DNA damage levels and tunes proliferative heterogeneity in cell populations[102]. This combined DDR network, the coordinated activity of p53, p16, p21, and SASP, forms the basis of the molecular etiology of skin photoaging. It connects DNA-damage sensing to cellular senescence and inflammatory remodeling and contributes to the phenotypic changes in the aging and UV-damaged skin[87,97,100].
3.5 Mitochondrial dysfunction and autophagy dysregulation
Mitochondrial dysfunction in photoaged skin results from cumulative mitochondrial DNA injury, impaired oxidative phosphorylation, altered mitochondrial dynamics, and reduced bioenergetic reserve. These disturbances decrease ATP production and increase ROS generation, thereby reinforcing oxidative stress, cellular senescence, and apoptosis[15,75,77].
Autophagic dysregulation is exemplified as a mismatch between autophagy activation and lysosomal degradative ability, which is often caused by impairments in mitochondrial fatty acid oxidation, which induce ATP depletion and lysosomal pH imbalances. These mutations inhibit the autophagic flux and acidification to support effective cargo elimination[103]. The mechanistic nexus is connected to the signaling cascades, such as AMP-activated protein kinase (AMPK), an energy sensor and autophagy induction regulator, and transcription factor EB (TFEB), the ultimate regulator of lysosomal biogenesis and autophagy gene transcription; these are both functionally impaired upon mitochondrial stress[15,104,105].
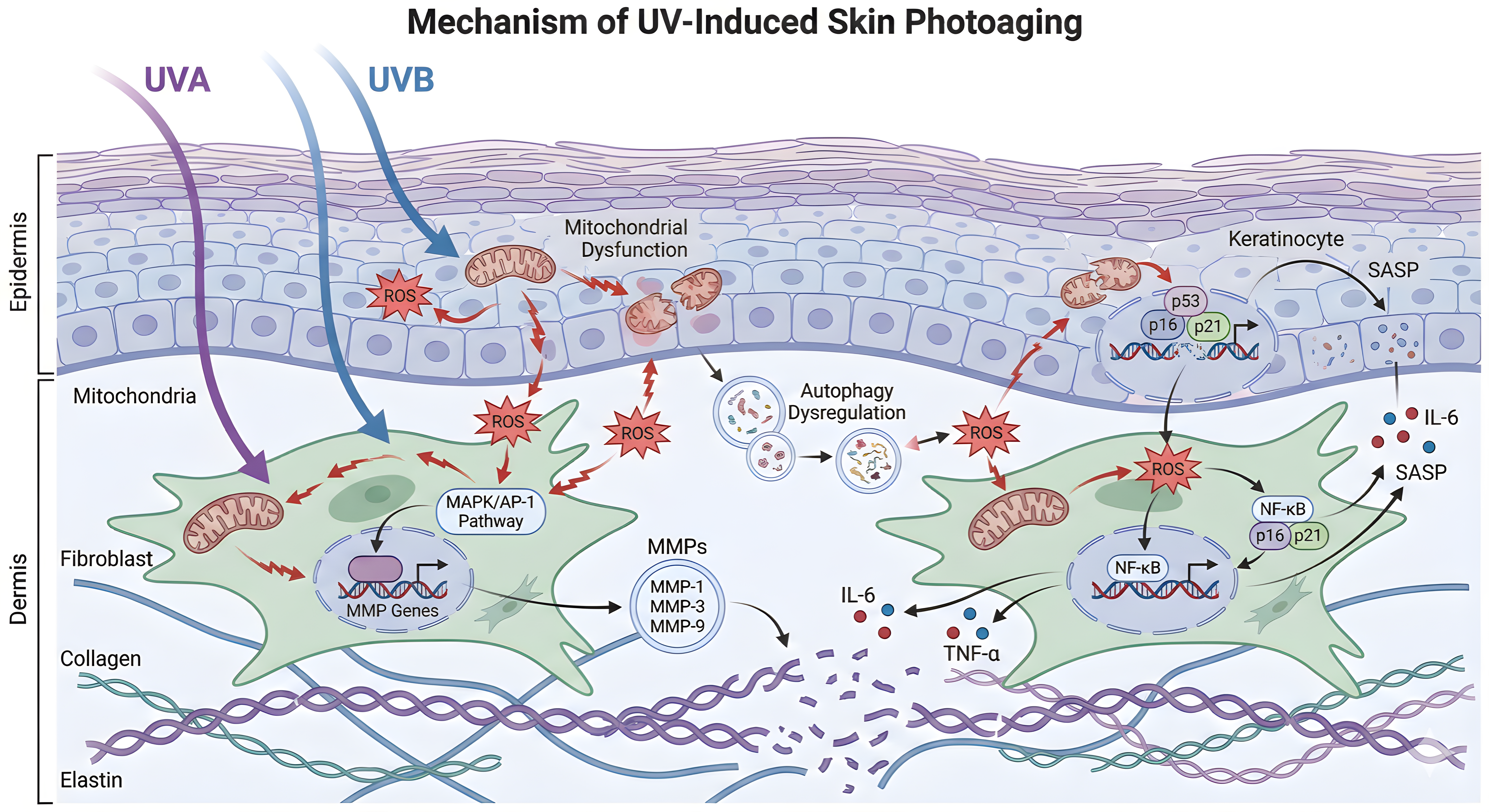
Together, mitochondrial impairment and defective autophagy create a feed-forward loop in which damaged mitochondria are inefficiently cleared, ROS production persists, and inflammatory and senescence pathways remain activated. Restoring mitophagy, lysosomal function, or energy-sensing pathways such as AMPK and TFEB may therefore represent therapeutic opportunities for attenuating photoaging-associated tissue decline[15,106,107]. The molecular mechanisms of UV-induced skin photoaging are illustrated in Figure 1 and summarized by pathway in Table 1.
{kind=link}
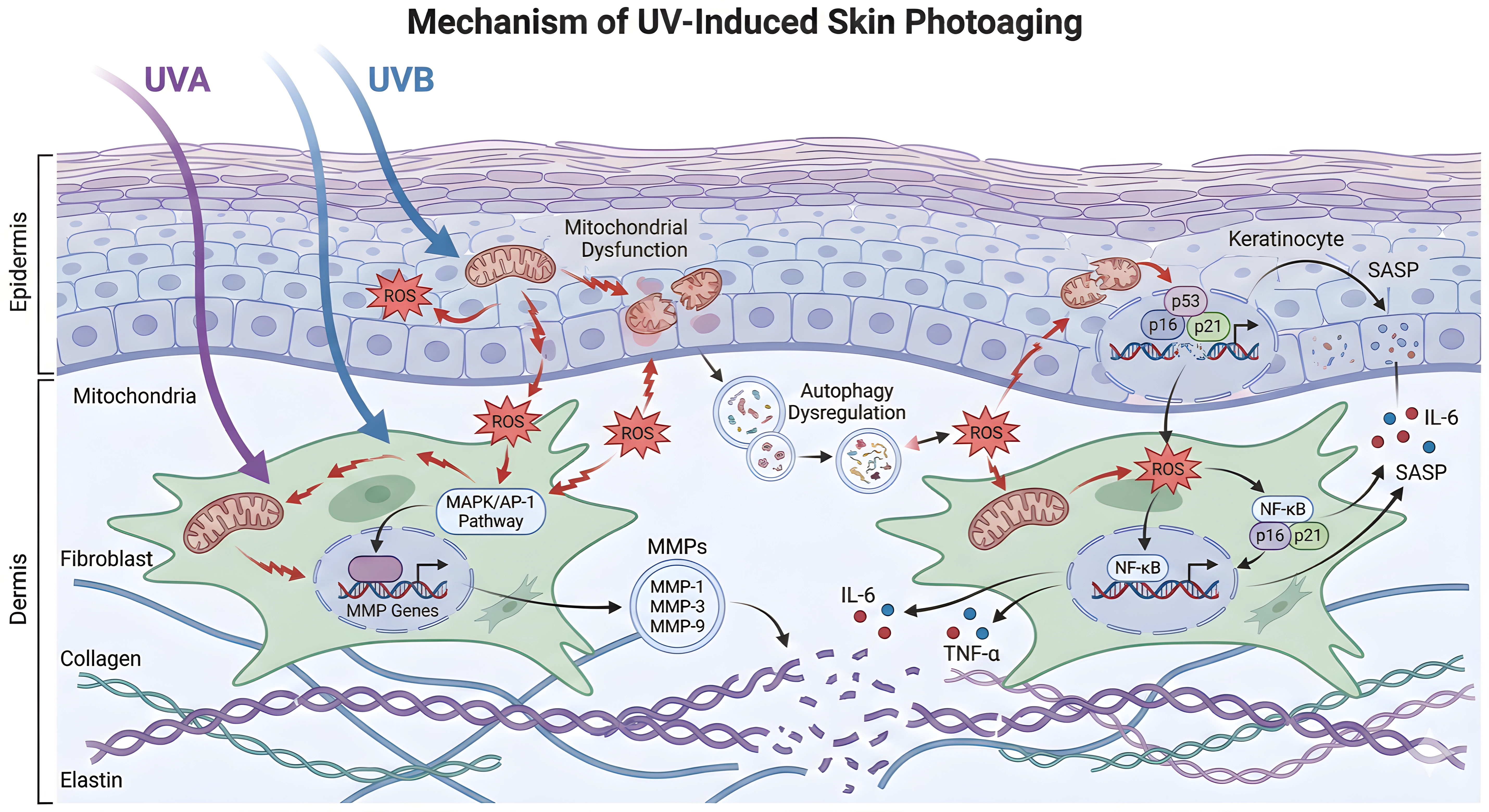
Figure 1. Molecular mechanisms of UV-induced skin photoaging. UVA: ultraviolet A; UVB: ultraviolet B; ROS: reactive oxygen species; MAPK: mitogen-activated protein kinase; AP-1: activator protein-1; MMPs: matrix metalloproteinases; ECM: extracellular matrix; NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells; IL-6: interleukin-6; TNF-α: tumor necrosis factor-alpha; SASP: senescence-associated secretory phenotype.
Table 1. Key molecular pathways in skin photoaging.
| Pathway/Event | Mechanism and Outcome | Citations |
| UV→ROS→Oxidative Stress | UV induces ROS, causing oxidative damage to DNA, proteins, and lipids | [15,72,108] |
| MAPK/AP-1→MMPs→ECM Degradation | ROS activates MAPK/AP-1, upregulates MMPs, degrades collagen/ECM | [72,79,81] |
| NF-κB→Cytokines | NF-κB induces IL-6, TNF-α, COX-2, sustaining inflammation and MMP expression | [74,88,89] |
| DNA Damage Response | p53, p16, p21 mediate cell cycle arrest, apoptosis, SASP, amplifying inflammation | [15,95,96] |
| Mitochondrial Dysfunction/Autophagy | UV damages mitochondria, impairs autophagy, increases ROS, promotes senescence | [15,106,107] |
ROS: reactive oxygen species; ECM: extracellular matrix; MAPK: mitogen-activated protein kinase; AP-1: activator protein-1; MMPs: matrix metalloproteinases; NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells; COX-2: cyclooxygenase-2; SASP: senescence-associated secretory phenotype.
4. Natural Extract-Based Interventions Against Skin Photoaging
4.1 Mechanistic rationale for natural extract-based photoprotection
Natural extract-based photoprotection is mechanistically plausible because many compounds target the same biological nodes that are disrupted by UV exposure. Across plant, marine, and microbiome sources, the most consistently reported mechanisms include ROS scavenging, activation of endogenous antioxidant defenses, suppression of MAPK/AP-1 and NF-κB signaling, inhibition of MMP-mediated ECM degradation, modulation of mitochondrial function, and reinforcement of epidermal barrier homeostasis.
4.2 Plant-derived bioactives
Plant-derived extracts are among the most extensively investigated natural interventions for UV-induced skin damage. Their activity is thought to be due to polyphenols, flavonoids, carotenoids, terpenoids, and other metabolites that help decrease ROS accumulation, inhibit inflammatory signaling, and maintain the integrity of the dermal extracellular matrix[109-111]. Thus, plant-based extracts are best regarded as adjunctive modulators of photoaging rather than stand-alone therapies[43,109].
Many plant extracts share the same mechanisms, despite their chemical diversity. Polyphenols and carotenoids attenuate oxidative stress in keratinocytes and fibroblasts, whereas several flavonoids and terpenoids suppress MAPK/AP-1 and NF-κB signaling, thereby reducing cytokine release and MMP-mediated collagen degradation[45,109,112]. The activation of the Nrf2/antioxidant response element (ARE) pathway is also crucial as it increases the intrinsic antioxidant capacity and facilitates cellular adaptation to the UV-induced stress[43,109,113].
These general mechanisms are exemplified by representative whole plant extracts that demonstrate their photoprotective effects. Polypodium leucotomos is a standardized extract of the fern that contains polyphenolic and organic acid constituents, including phenolic acids like ferulic, caffeic and vanillic acids[114]. It increases antioxidant defense and decreases oxidative stress and inflammation induced by UV and high-energy visible light (HEVL) such as ROS generation and TNF-α secretion and promotes the maintenance of immune homeostasis in the skin[115-117]. PLE also blocks the formation of dark-CPD following UVA exposure[118] and suppresses melanin production in melanocytes in vitro after exposure to HEVL[119]. Clinically, it enhances the minimal erythema dose and diminishes erythema caused by UVB radiation[120-122]. Reseda luteola contains a high amount of luteolin, which has anti-oxidative, anti-erythema, and anti-pigmentation effects by inhibiting NF-κB, MAPK, and AP-1 signaling and reducing MMP-1 and COX-2 expression[117,123,124]. Licorice (Glycyrrhiza spp.) extracts contain several flavonoids and chalcones, with licochalcone A being a key bioactive compound that is able to activate the Nrf2 signaling pathway, reduce oxidative stress, and prevent HEVL-induced pigmentation in vitro and in vivo[117,125,126].
Other botanicals are also worth considering, including Hypericum perforatum, which contains hyperforin-rich fractions that exhibit antioxidant and anti-inflammatory effects and have potential for Nrf2-mediated cytoprotection but also hypericin, which can cause photosensitization; this underscores the importance of careful formulation[88,117,127]. Potentilla erecta is rich in bioactive ellagitannins, flavonoids (quercetin) and phenolic acids, which are responsible for its anti-photoaging activity against erythema, pigmentation and DNA damage[117,128]. Deschampsia antarctica aqueous extract (commercially available as Edafence®) involves reducing oxidative pigmentation in melanocytes by blocking HEVL-induced melanogenesis[117,129].
The mechanistic basis of these photoprotective effects becomes clearer when examining individual bioactive phytochemicals isolated from plant extracts. Resveratrol, a stilbene polyphenol found in grape skin and other plants, protects against UV-induced photoaging by reducing ROS accumulation, suppressing apoptosis, and activating AMPK-dependent autophagy. In experimental models, these effects translated into reduced MMP-1 expression, preservation of collagen integrity, and improvement of photoaging-associated phenotypes including wrinkles, erythema, hyperpigmentation, and inflammation[45,130]. The principal compound in green tea (Camellia sinensis), epigallocatechin-3-gallate (EGCG), downregulates ROS, MDA, and 8-OHdG, restores antioxidants, preserves telomere integrity, normalizes cell-cycle regulation, and inhibits p38 MAPK, NF-κB, and AP-1 signaling, thereby reducing MMP-1 and inflammatory cytokines[76,112,131].
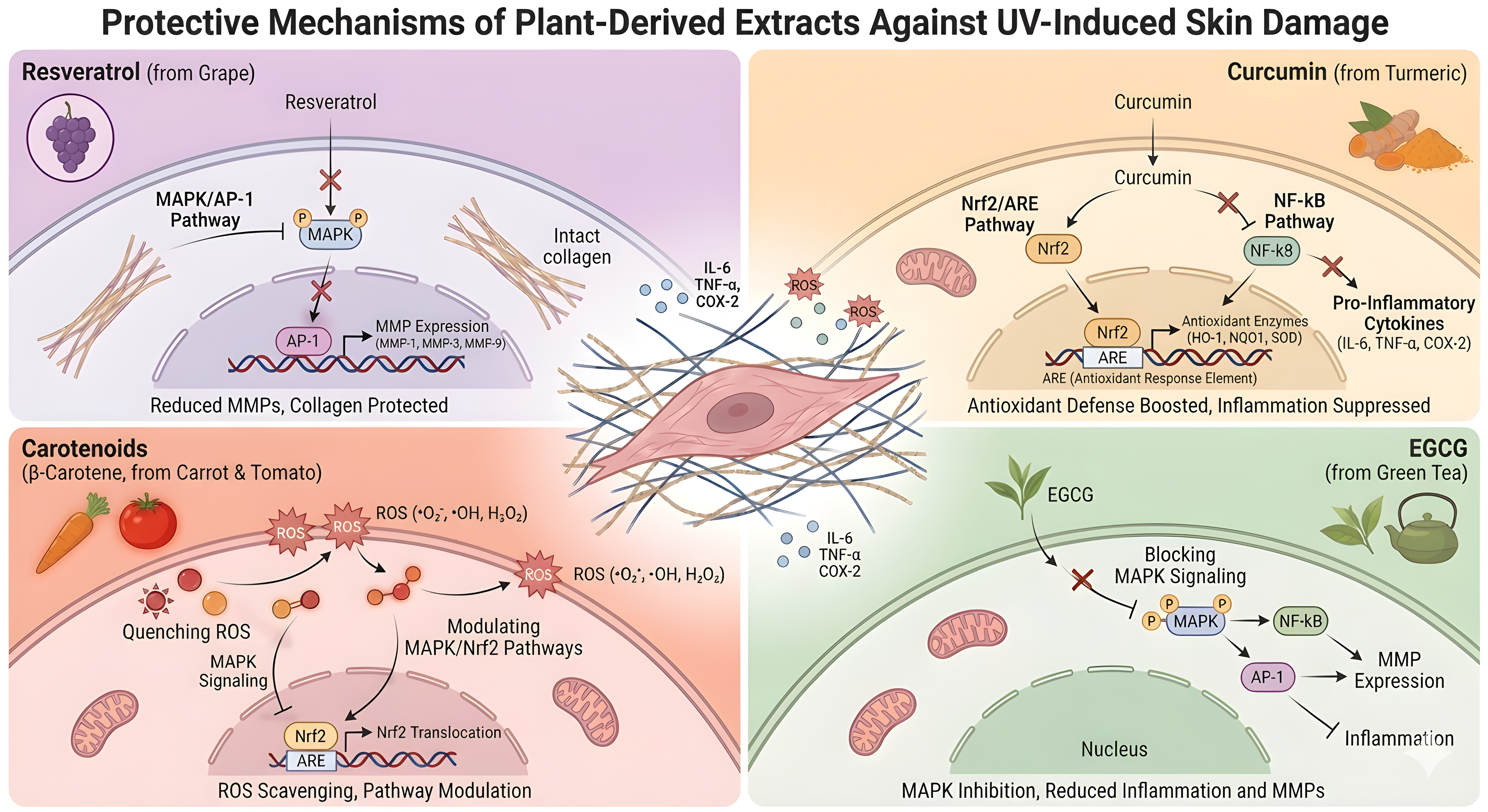
The major curcuminoid of turmeric (Curcuma longa), curcumin, protects against UVB-induced skin damage by decreasing ROS and apoptosis, enhancing mitochondrial function, inducing mitochondrial autophagy via YAP1 signaling, and inhibiting NLRP3/IL-18 inflammatory signaling in UVB-irradiated mouse skin and HaCaT keratinocytes[132]. In line with this, the expression of TNF-α was also decreased and the expression of type I collagen was increased in mouse skin exposed to UVB and treated with turmeric extract containing curcumin[133]. Carotenoids are another popular plant-based photoprotection ingredient. A randomized clinical study showed that UV-induced erythema decreased and serum β-carotene increased in humans after supplementation with banana pulp-derived β-carotene, leading to systemic photoprotection[134]. In parallel, tomato carotenoids have been shown to inhibit NF-κB activation and IL-6 production in keratinocytes, to act synergistically with Nrf2/ARE activation, and to reduce MMP-1 signaling in fibroblasts, indicating that carotenoid-containing formulations can affect both oxidative stress and inflammatory pathways related to photoaging[111]. Other compounds, such as ginsenosides, proanthocyanidins, and quercetin, exhibit anti-apoptotic, anti-inflammatory, and ECM-preserving effects[43,135,136]. The summary of the representative plant-derived natural extracts’ protective mechanisms is shown in Figure 2.
{kind=link}
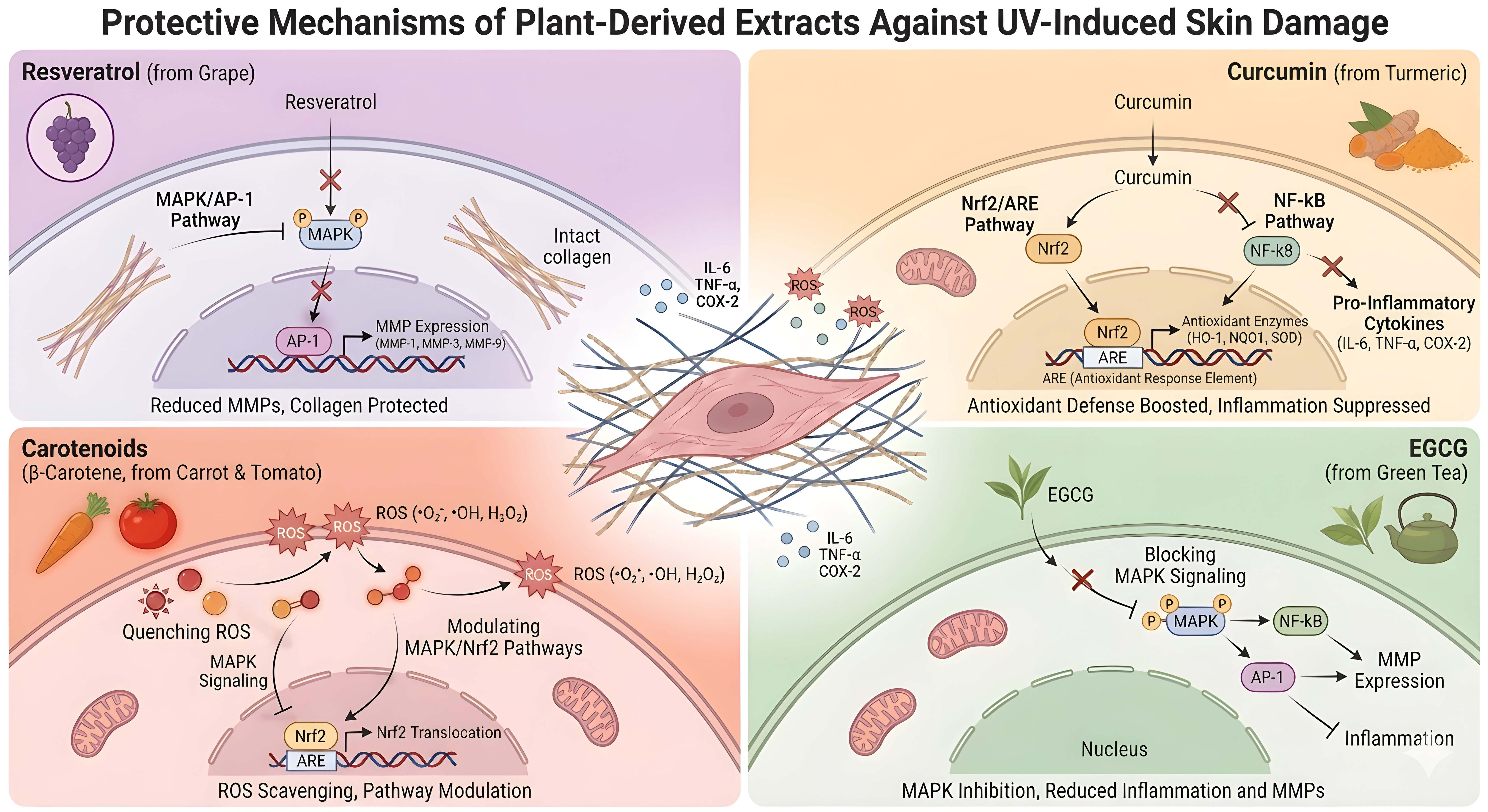
Figure 2. Protective mechanisms of representative plant-derived extracts against UV-induced skin damage. UV: ultraviolet; ROS: reactive oxygen species; MAPK: mitogen-activated protein kinase; AP-1: activator protein-1; MMPs: matrix metalloproteinases; ECM: extracellular matrix; NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells; Nrf2: nuclear factor erythroid 2-related factor 2; ARE: antioxidant response element; EGCG: epigallocatechin gallate.
In recent years, macromolecules and vesicles derived from plants have been the subject of attention. These are polysaccharides, proteins, and extracellular vesicle-like nanostructures that are used as carriers or stabilizers to enhance the stability, penetration, and bioavailability of active compounds without adding new pharmacology[43,113,137]. In general, plant extracts are supportive antioxidants and anti-inflammatory agents in well-defined UV-damage pathways. They inhibit the degradation of collagen and elastin, chronic inflammation and progressive photoaging by scavenging ROS, upregulating Nrf2/ARE and suppressing AP-1/NF-κB signaling[109,113,138].
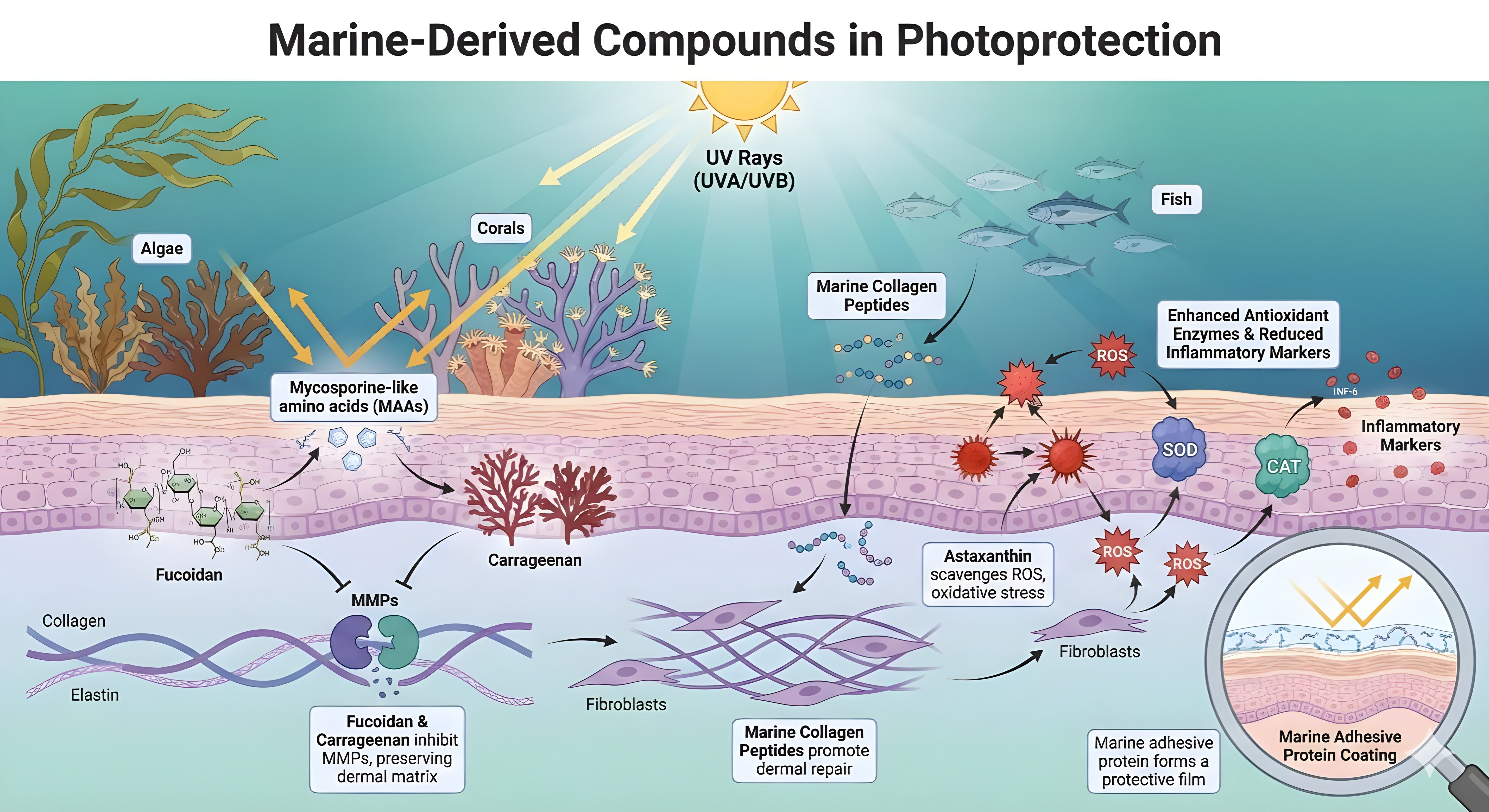
4.3 Marine-derived bioactives
Marine-derived compounds contribute unique mechanisms to photoprotection that complement terrestrial plant extracts. Mycosporine-like amino acids (MAAs), produced by algae and coral-associated organisms, absorb UV photons and dissipate the energy as heat. This physical shielding reduces DNA strand breaks and cyclobutane pyrimidine dimer formation in keratinocytes[139-141]. Chitosan nanoformulations of MAA-rich red algal extracts also provide strong UVA photoprotection in HaCaT keratinocytes and preserve viability at high UVA doses[139].
Sulfated polysaccharides, such as fucoidan and carrageenan, provide downstream protection by inhibiting matrix metalloproteinases (MMP-1, MMP-3, MMP-9). These macromolecules act as competitive inhibitors of elastase and collagenase, helping preserve collagen type I and elastin fibers in the dermal extracellular matrix[140-142]. Together, these upstream (MAAs) and downstream (polysaccharides) actions form a dual-mechanistic paradigm that robustly protects against UV-induced matrix degradation[139,142].
The control of oxidative stress and the regulation of inflammatory cascades are the key elements of preventing the UV-induced damage. The conceptual diagram highlights the central role of marine xanthophylls, in particular, astaxanthin in neutralizing reactive oxygen species produced by fibroblasts[142,143]. Due to its special molecular structure with a polar-nonpolar-polar configuration, astaxanthin penetrates the cell membrane and quenches singlet oxygen more effectively than β-carotene or tocopherol[144,145]. This full scavenging is accompanied by the upregulation of endogenous antioxidant enzymes, such as Superoxide Dismutase and Catalase[142,146].
In parallel, marine bioactives inhibit pro-inflammatory mediators, interrupting the chronic inflammatory loop that drives dermal senescence and maintaining fibroblast viability under oxidative stress[142,143]. The summary of the function and mechanism of marine-derived natural extracts as shown in Figure 3.
{kind=link}
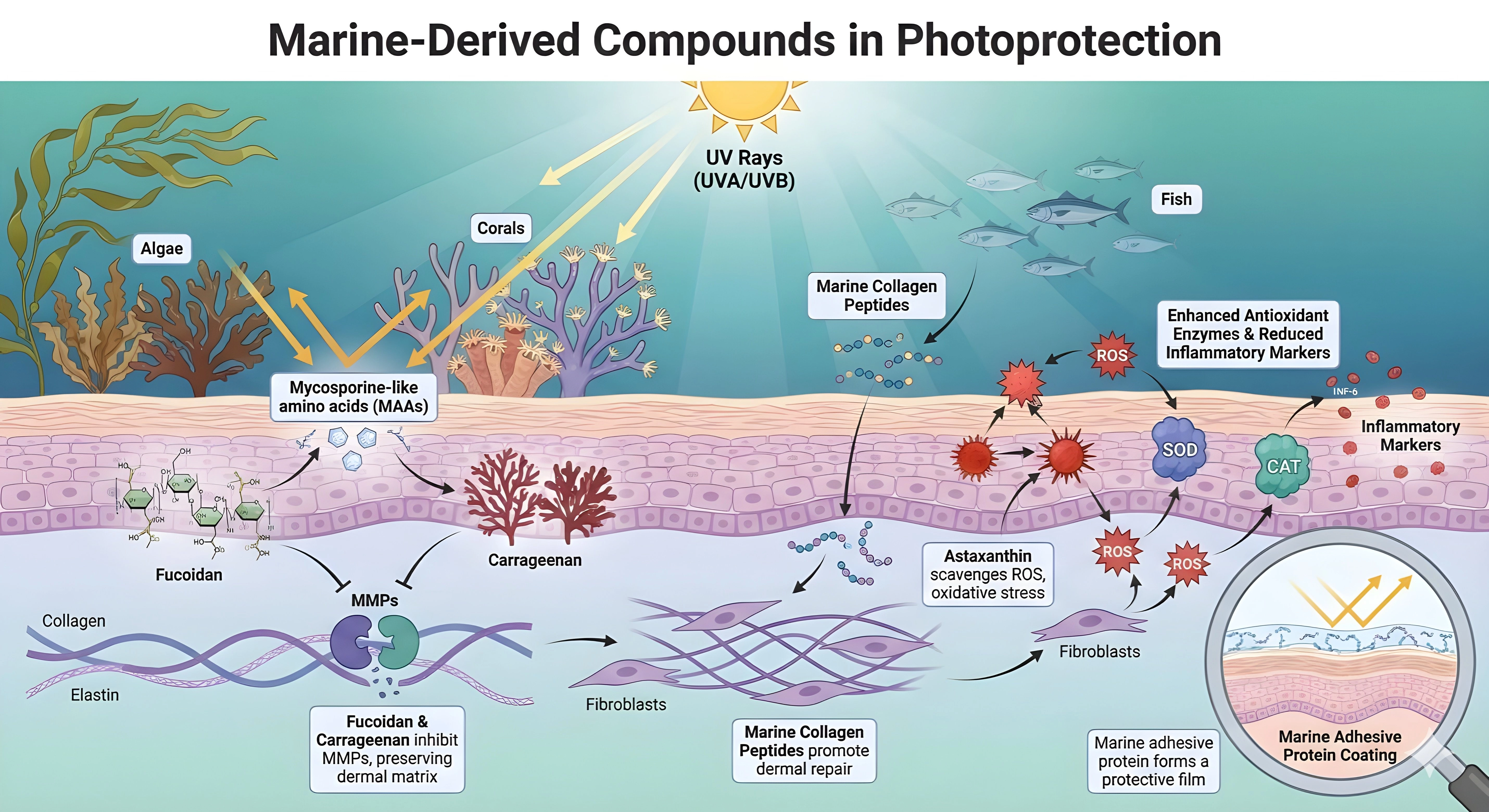
Figure 3. Marine-derived compounds in photoprotection. MAAs: mycosporine-like amino acids; ROS: reactive oxygen species; MMPs: matrix metalloproteinases; ECM: extracellular matrix; SOD: superoxide dismutase; CAT: catalase.
Beyond photoprotection and antioxidant defense, marine compounds support structural restoration. Collagen peptides penetrate the dermis, where they stimulate fibroblast proliferation and de novo pro-collagen synthesis[147]. Low-molecular-weight hydrolysates enhance skin hydration and viscoelasticity by increasing hyaluronic acid production[147,148]. Marine adhesive proteins form a biocompatible extracellular coating that aids wound healing and re-epithelialization, completing a multi-level photoprotective mechanism encompassing UV shielding, matrix preservation, and barrier reinforcement[149,150].
4.4 Microbiome- and probiotic-derived interventions
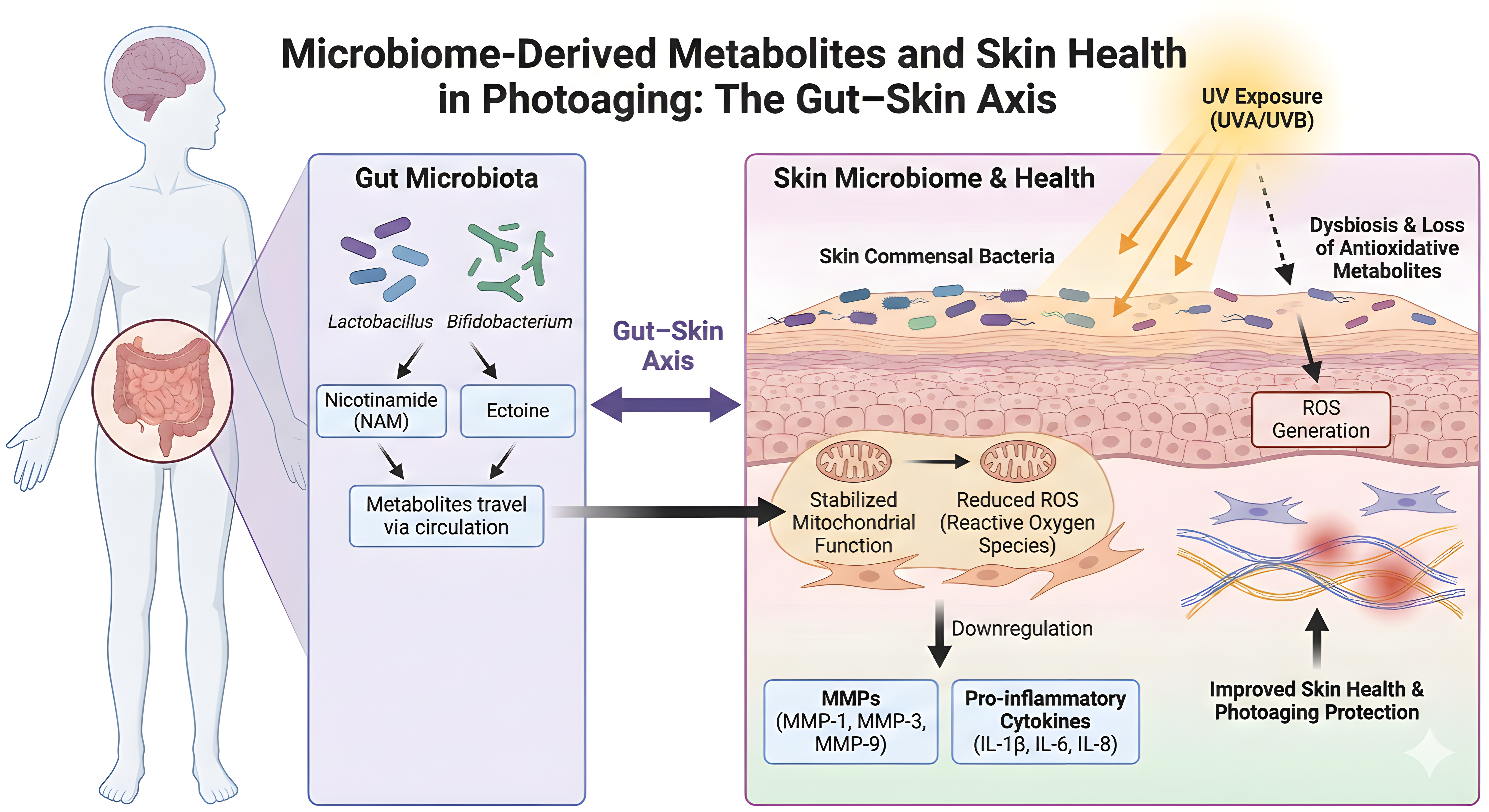
Recent studies of the gut–skin axis have revealed a systemic pathway through which intestinal microbiota mitigate cutaneous photodamage. Commensal bacteria, particularly species of Lactobacillus and Bifidobacterium, act as bio-factories of protective metabolites such as nicotinamide (NAM) and ectoine[151,152]. Selected Lactobacillus and Bifidobacterium strains improve UV injury phenotypes and support skin–gut communication[153-155]. In particular, Limosilactobacillus fermentum XJC60-derived NAM lowers ROS, stabilizes mitochondrial membrane potential, restores the NAD+/NADH ratio, and downregulates MMP-1, MMP-3, and inflammatory cytokines, with preservation of collagen fibers in vivo[156]. Unlike topical agents, these bioactive molecules are absorbed into the bloodstream and delivered to the dermis, providing an “internal sunscreen” effect[157-159].
Metagenomic and metabolomic analyses show that oral administration of these probiotics significantly alters the serum metabolome, increasing the bioavailability of short-chain fatty acids and NAD+ precursors, which are crucial for maintaining skin homeostasis[160,161]. This systemic route also addresses dysbiosis of the skin microbiome caused by acute UV exposure, thereby restoring microbial barrier function[162,163].
At the cellular level, the protective effects of microbiome-derived metabolites are mediated primarily through mitochondrial stabilization and regulation of oxidative stress. UV radiation increases ROS, causing mitochondrial dysfunction in keratinocytes and fibroblasts (Figure 4)[164]. Microbiota-associated metabolites such as NAM act as antioxidants and bioenergetic regulators, maintaining mitochondrial membrane potential and upregulating endogenous antioxidant enzymes[154,156,165]. These effects interrupt the feed-forward loop of ROS amplification, preventing downstream activation of senescence pathways and preserving dermal cell viability[153,166,167].
{kind=link}
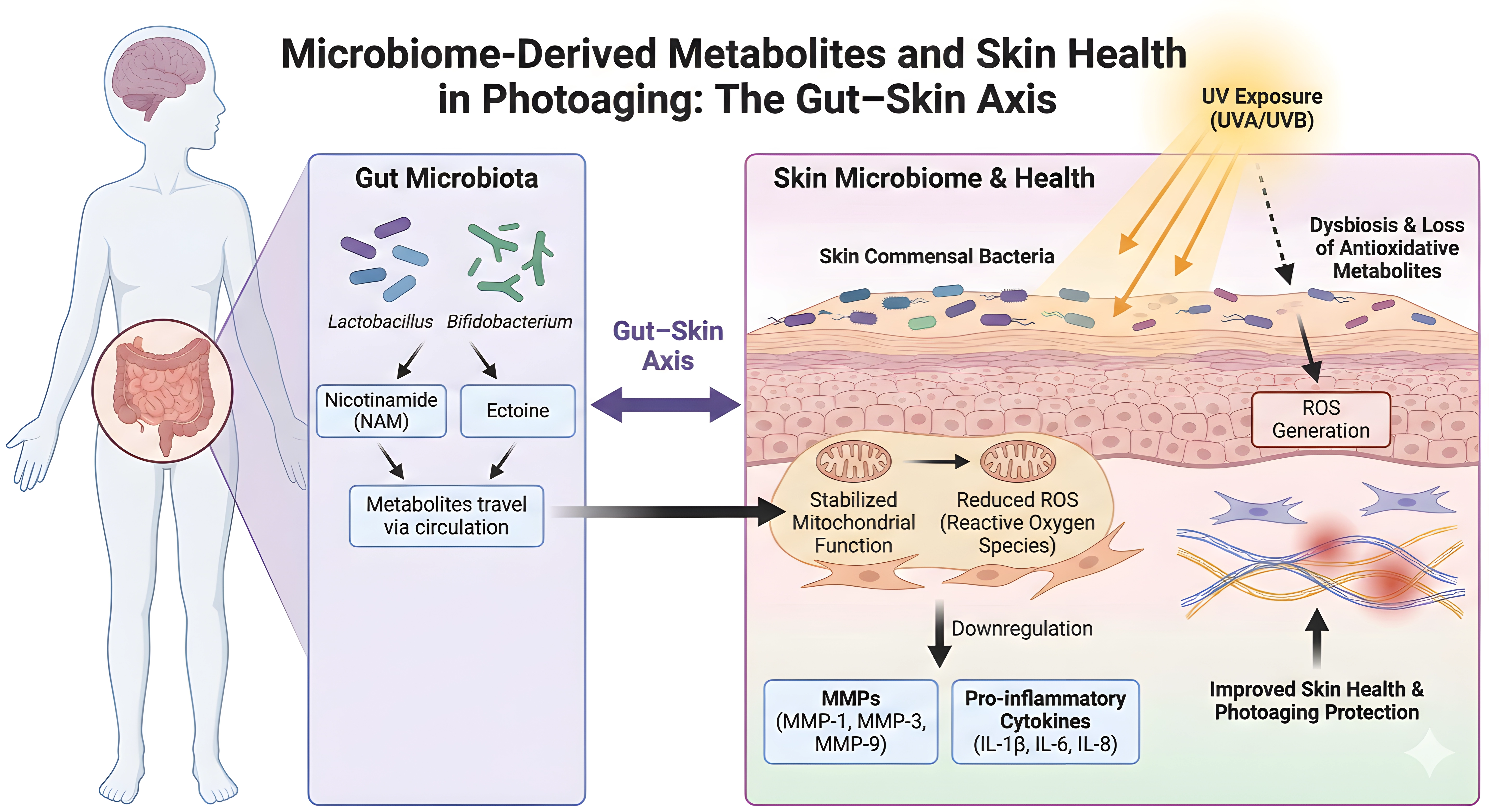
Figure 4. Microbiome-derived metabolites and skin health in photoaging: the gut-skin axis. NAM: nicotinamide; ROS: reactive oxygen species; MMPs: matrix metalloproteinases; IL: interleukin; TNF-α: tumor necrosis factor-alpha; NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells; ECM: extracellular matrix.
Structurally, this gut–skin signaling confers additional benefits by inhibiting inflammation and preserving the ECM. As illustrated in (Figure 4), probiotic-derived metabolites and live or heat-killed strains downregulate MMPs (MMP-1, MMP-3, MMP-9) and pro-inflammatory cytokines, limiting ECM degradation[153,155,156]. In vivo studies confirm that probiotic supplementation suppresses AP-1 signaling, reducing MMP transcription and consequent cleavage of collagen and elastin fibers[153,154]. Furthermore, gut-derived metabolites enhance skin hydration and decrease wrinkle depth[153-155], while their broader anti-inflammatory effects may also help constrain NF-κB-driven chronic inflammatory signaling[168,169].
4.5 Comparative and synergistic strategies
Natural interventions derived from plant, marine, and microbiome sources have distinct mechanisms of action but share common anti-photoaging endpoints. Extracts of these plants are rich in polyphenols, flavonoids, and carotenoids, which primarily modulate intracellular redox balance and inflammatory signaling. They work through important pathways, including Nrf2/ARE and MAPK/AP-1, decreasing oxidative stress, MMP activity, and pro-inflammatory cytokine release, and related PI3K/Akt signaling remains of interest[45,109,110].
Marine extracts, especially from seaweeds, contain UV-absorbing compounds and ECM protectants, such as MAAs, and chitosan nanoformulations of MAA-rich red algal extracts have been demonstrated to protect HaCaT keratinocytes from UVA damage[139]. These compounds have been demonstrated to have ROS-scavenging properties, to inhibit MMPs, and to protect keratinocytes from UV-induced DNA damage physically. Several of these effects have been proven in human trials and include smoother skin and photoprotection[4,170].
Microbiome-derived interventions, notably probiotics such as Limosilactobacillus fermentum, help to maintain immune-metabolic homeostasis and strengthen barrier function. In UV injury models, L. fermentum XJC60-derived nicotinamide reduces ROS, maintains the MMP, replenishes the NAD+/NADH ratio, and downregulates MMP-1, MMP-3, and inflammatory cytokines while maintaining collagen fibers[156]. L. plantarum HY7714 exopolysaccharide protects against skin aging through skin–gut axis communication[165], and Bifidobacterium animalis subsp. lactis MG741 as well as L. fermentum MG5368/Lactiplantibacillus plantarum MG989 reduce MMPs, AP-1/MAPK signaling, and wrinkle-related outcomes in UVB-exposed cells and mice[153,154]. These interventions also interact with the host micro-environment to restore microbial balance disrupted by UV exposure[1,22,156].
Rational combination strategies may maximize photoprotective efficacy. Pairing a ROS-scavenging botanical com-pound with a marine-derived MMP inhibitor, a barrier-supportive peptide, and a probiotic or postbiotic metabolite could address the multi-layered nature of skin photoaging. These combinations should be evaluated using standardized endpoints, including TEWL, erythema, pigmentation, wrinkle depth, collagen density, MMP expression, cytokine pro-files, and microbiome composition[22,127,170]. This integrated approach aligns with a mechanism-driven paradigm, wherein distinct interventions target complementary pathways while converging on shared structural and functional outcomes in photoaged skin (Figure 5, Table 2).
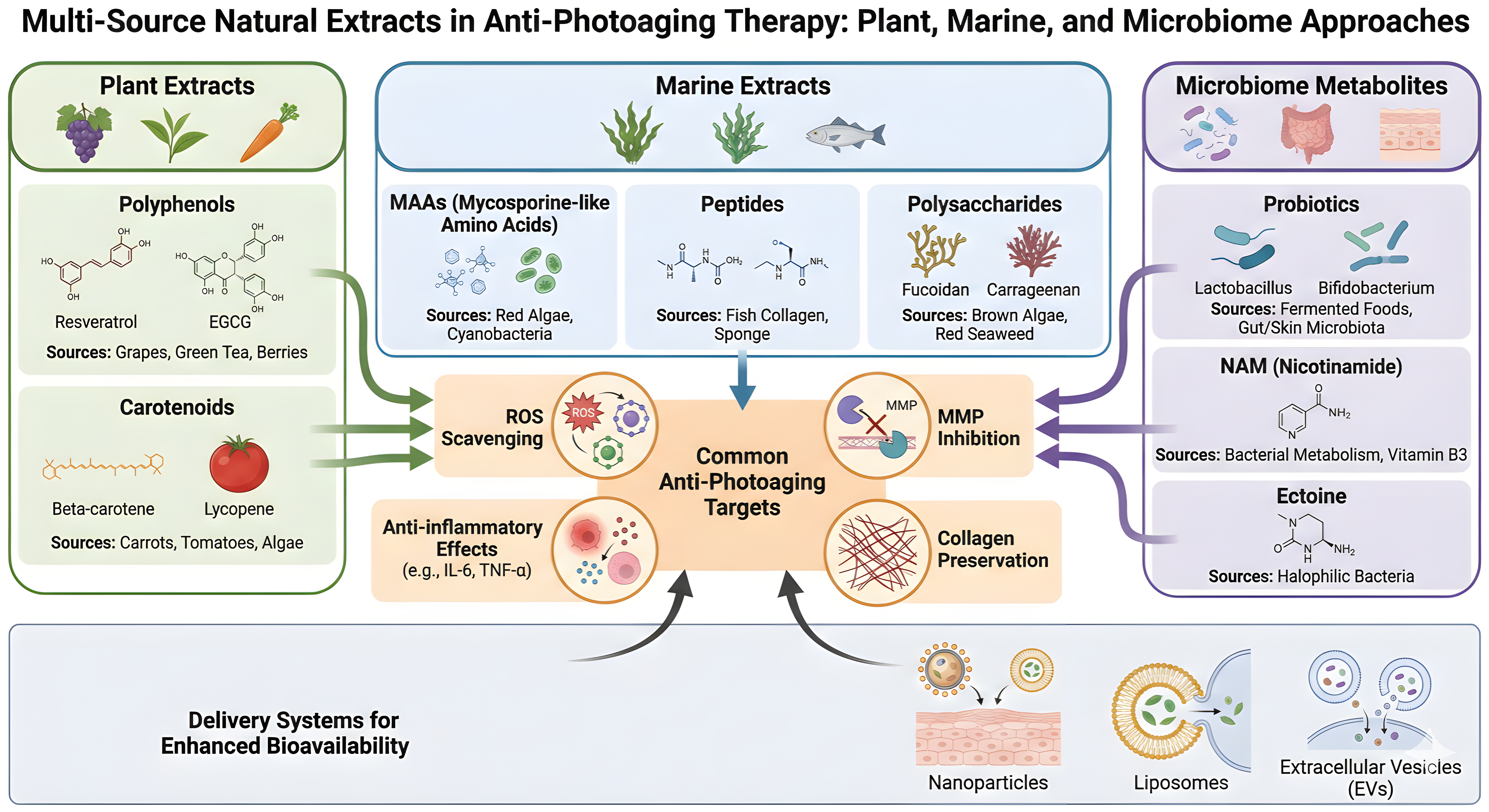{kind=link}
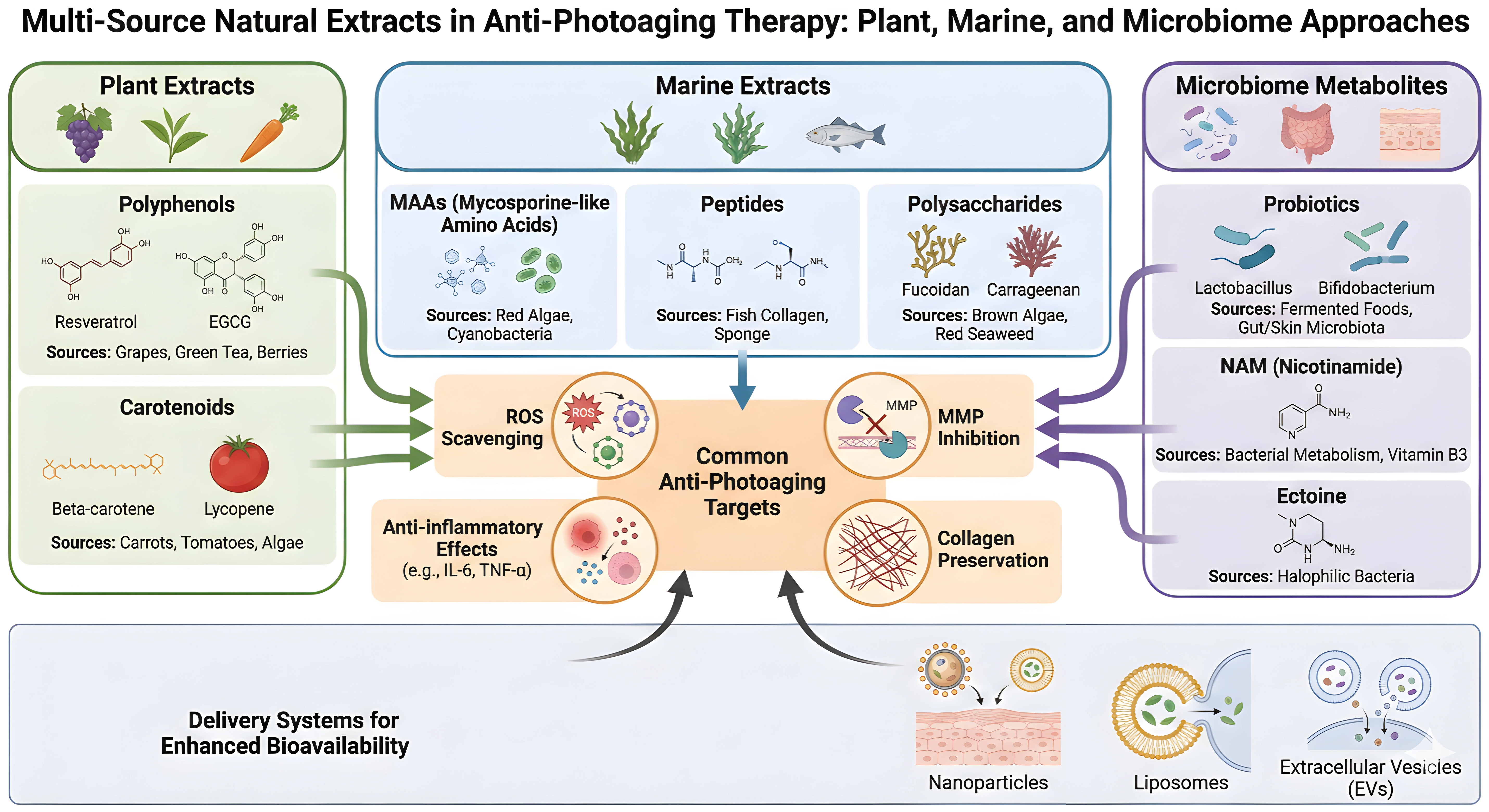
Figure 5. Multi-source natural extracts in anti-photoaging therapy: plant, marine, and microbiome approaches. EVs: extracellular vesicles; MAAs: mycosporine-like amino acids; ROS: reactive oxygen species; MMPs: matrix metalloproteinases; NAM: nicotinamide; ECM: extracellular matrix.
Table 2. Comparison of plant, marine, and microbial extracts for skin photoaging: targets, efficacy, and translational potential.
| Extract Source | Key Molecular Targets | Efficacy Highlights | Translational Potential | Citations |
| Plants | Nrf2, MAPK, PI3K/Akt, MMPs | Antioxidant, anti-inflammatory, DNA repair, skin barrier repair | High (clinical/animal data, advanced delivery systems) | [43,45,109,110] |
| Marine Organisms | ROS, MMPs, UV absorption | Potent UV protection, MMP inhibition, clinical validation | Moderate-High (unique actives, some clinical use) | [139,142,171,172] |
| Microbiome/Probiotics | Mitochondrial function, cytokines | ROS reduction, collagen preservation, immune modulation | Emerging (preclinical, potential for synergy) | [153-156] |
Nrf2: nuclear factor erythroid 2-related factor 2; MAPK: mitogen-activated protein kinase; PI3K/Akt: phosphoinositide 3-kinase/protein kinase B; MMPs: matrix metalloproteinases; ROS: reactive oxygen species; UV: ultraviolet.
5. Conclusion
Skin photoaging is a layer-specific and pathway-interconnected process rather than a simple outcome of oxidative injury. UVB is mainly responsible for inducing DNA photolesions, disrupting barrier function, and activating inflammatory signaling in the epidermis. At the same time, UVA penetrates deeper into the dermis, leading to ROS accumulation, mitochondrial dysfunction, fibroblast senescence, and ECM degradation. Subcutaneous adipose dysfunction also con-tributes to biomechanical deterioration and visible skin laxity. Recent evidence indicates that blue light increases ROS production and triggers melanogenesis via the Opsin-3 receptor, leading to long-lasting hyperpigmentation, especially in darker skin types[16]. Additionally, excess iron released from ferritin after UVA exposure leads to ROS generation and activation of MMP-1, which accelerates the degradation of dermal collagen[53,54].
Plant, marine, and microbiome natural extracts target several anti-photoaging mechanisms. The main intracellular tar-gets of plant-derived compounds include intracellular redox balance, inflammatory pathways, Nrf2/ARE signaling, MAPK/AP-1 signaling, and NF-κB signaling. Marine molecules such as mycosporine-like amino acids, polysaccharides and marine peptides, protect against UV radiation, ROS, ECM, and collagen. Microbiome-derived interventions have systemic effects by stabilizing mitochondria, decreasing ROS, regulating cytokine expression, and maintaining immune-metabolic homeostasis. These multi-source interventions all aim to achieve common goals, including ROS attenuation, MMP inhibition, collagen maintenance, barrier repair, and reducing chronic dermal inflammation.
The integrative synthesis highlights the translational potential of natural extracts as adjunctive photoprotective agents. They need to be standardized in their characterization, reproducible in their formulation, deliverable through the skin, and safe for long-term use. This review connects the dots between tissue-layer responses to UV exposure and the fundamental molecular mechanisms and natural interventions from multiple sources, thereby offering a framework for mechanism-driven photoprotection for guiding research and clinical practice.
6. Challenges and Future Perspectives
Although significant efforts have been made to elucidate the molecular mechanisms of skin photoaging and the therapeutic potential of natural extract-based interventions, several scientific and translational challenges remain. Extracts from plants, marine sources, and the microbiome are chemically complex. They can exhibit significant batch-to-batch variation, which can hinder the reproducibility and standardization of preclinical and clinical studies[127,173,174]. Moreover, many bioactive compounds, such as polyphenols, carotenoids, and peptides, have poor solubility and limited stability, poor skin penetration, and rapid metabolic degradation, resulting in poor bioavailability and in vivo efficacy[127,175,176]. In addition, photoaging is a multifactorial process that includes oxidative stress, mitochondrial dysfunction, inflammation, ECM remodeling, impaired autophagy, cellular senescence, and microbiome dysbiosis, and requires integrated approaches that can tackle multiple inter-connected pathways simultaneously[36,43]. Many potential interventions have shown promising photoprotective effects in in vitro or animal models. However, there is still a lack of strong human clinical evidence for many of these interventions[173,177,178].
Future studies are needed to standardize experimental and translational strategies. This includes controlled UVA/UVB and blue light exposure models, rigorous extract characterization (e.g., chemical fingerprinting and batch-to-batch quality control), and dose-response testing. Clinically meaningful endpoints, such as TEWL, erythema index, pigmentation, wrinkle depth, collagen density, MMP and cytokine profiling, and skin microbiome composition, should be measured to ensure translational relevance[173,179,180]. Furthermore, multi-omics analysis that combines transcriptomic, proteomic, metabolomic, and skin-phenotype data might offer more insight regarding host–microbiome–ECM interactions and help to identify optimized intervention strategies[181,182].
Mechanism-based strategies targeting key molecular pathways involved in photoaging should be increasingly used in future therapeutic development. The key molecular targets are the Nrf2/ARE axis for antioxidant defence[127,183], MAPK/AP-1 and NF-κB signalling pathways for inflammation and MMP regulation[45,177], TGF-β/Smad signalling pathways for collagen synthesis and ECM maintenance[180], and AMPK/SIRT1 and mTOR/autophagy pathways for mitochondrial and cellular homeostasis[130,184]. Other targets include senescence-associated proteins such as p53/p21 and p16INK4a/Rb, and proteins associated with epidermal barrier integrity, such as filaggrin and tight junction components[127]. Combining multiple pathways in a single intervention using natural compounds may be more effective than single-pathway interventions. Specifically, plant polyphenols (e.g., resveratrol), marine peptides (e.g., collagen hydrolysates), carotenoids (e.g., astaxanthin), and microbiome-derived metabolites may exhibit synergistic effects by targeting multiple photoaging hallmarks simultaneously[176,178,185].
Sustainability, safety, and regulatory compliance will be equally important for the successful translation of these interventions into clinical and commercial applications. The use of agrifood waste streams as sources of bioactive ingredients, including citrus peels, is in line with the principles of the circular economy and helps to minimize environmental impact[186,187]. Likewise, marine bioactives, such as collagen peptides, should be obtained from sustainable aquaculture or biotechnological production systems to reduce the ecological impact[176]. Regulatory frameworks should also include extract standardization, batch consistency, cytotoxicity, allergenicity, contamination control, label accuracy, and safety validation using high quality primary evidence[174]. Future studies involving interdisciplinary collaboration among molecular biologists, dermatologists, microbiologists, and formulation scientists can help develop safe, effective, and mechanism-based natural interventions for the prevention and management of skin photoaging.
Acknowledgement
The authors acknowledge the use of AI-assisted tools, Grammarly, for language editing and improvement of clarity, and Gemini (Google) for assistance in figure generation. All content, figures, and interpretations were reviewed and verified by the authors, who take full responsibility for the accuracy and integrity of the work.
Authors contribution
Lai J, Ren X: Investigation, data curation, writing-original draft.
Liang G, Jia S, Li J, Wang Y, Zhang H: Validation, writing-review & editing.
Xie W: Supervision, conceptualization, project administration, writing-review & editing.
All authors have read and agreed to the published version of the manuscript.
Conflicts of interest
Weidong Xie is an Editorial Board Member of Ageing and Cancer Research & Treatment. Gaobin Liang is employed by Guangzhou Hejin Biotechnology Co., Ltd. This study was supported by a cooperation project between Tsinghua Shenzhen International Graduate School and Guangzhou Hejin Biotechnology Co., Ltd. The other authors declare no conflicts of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Funding
This work was supported by the cooperation project of Tsinghua Shenzhen International Graduate School and Guangzhou Hejin Biotechnology Co., Ltd. [Grant No. 20229660163].
Copyright
© The Author(s) 2026.
References
-
9. Kaltchenko MV, Chien AL. Photoaging: Current concepts on molecular mechanisms, prevention, and treatment. Am J Clin Dermatol. 2025;26(3):321-344.[DOI]
-
13. Ichihashi M, Ueda M, Budiyanto A, Bito T, Oka M, Fukunaga M, et al. UV-induced skin damage. Toxicology. 2003;189(1-2):21-39.[DOI]
-
14. De Los Santos Gomez P, Costello L, Goncalves K, Przyborski S. Comparison of photodamage in non-pigmented and pigmented human skin equivalents exposed to repeated ultraviolet radiation to investigate the role of melanocytes in skin photoprotection. Front Med. 2024;11:1355799.
-
23. Lv M, Yang Y, Choisy P, Xu T, Pays K, Zhang L, et al. Flavonoid components and anti-photoaging activity of flower extracts from six Paeonia cultivars. Ind Crops Prod. 2023;200:116707.[DOI]
-
25. Sharma P, Dhiman T, Negi RS, Anshad OC, Gupta K, Bhatti JS, et al. A comprehensive review of the molecular mechanisms driving skin photoaging and the recent advances in therapeutic interventions involving natural polyphenols. S Afr N J Bot. 2024;166:466-482.[DOI]
-
35. Fan Z, Zhou Y, Gan B, Li Y, Chen H, Peng X, et al. Collagen-EGCG combination synergistically prevents UVB-induced skin photoaging in nude mice. Macromol Biosci. 2023;23(12):2300251.[DOI]
-
39. Mayangsari E, Mustika A, Nurdiana N, Samad N. Comparison of UVA vs UVB photoaging rat models in short-term exposure. Med Arch. 2024;78(2):88.[DOI]
-
41. Kajitani GS, Quayle C, Garcia CCM, Fotoran WL, dos Santos JFR, van der Horst GTJ, et al. Photorepair of either CPD or 6-4PP DNA lesions in basal keratinocytes attenuates ultraviolet-induced skin effects in nucleotide excision repair deficient mice. Front Immunol. 2022;13:800606.
-
45. Cui B, Wang Y, Jin J, Yang Z, Guo R, Li X, et al. Resveratrol treats UVB-induced photoaging by anti-MMP expression, through anti-inflammatory, antioxidant, and antiapoptotic properties, and treats photoaging by upregulating VEGF-B expression. Oxid Med Cell Longev. 2022;2022:6037303.
-
48. Lohakul J, Chaiprasongsuk A, Jeayeng S, Saelim M, Muanjumpon P, Thanachaiphiwat S, et al. The protective effect of polyherbal formulation, harak formula, on UVA-induced photoaging of human dermal fibroblasts and mouse skin via promoting Nrf2-regulated antioxidant defense. Front Pharmacol. 2021;12:649820.
-
56. Bighetti S, Rovati C, Bettolini L, Arisi M, Rossi M, Ravelli C, et al. Two-photon microscopy for the investigation of morphological and quantitative changes in skin chrono- and photo-aging. Cosmetics. 2025;12(3):111.[DOI]
-
58. Ma Y, Huang J, Gong J, Li L, Zhao Y, Jin Y, et al. Colla Corii Asini regulate collagen regeneration in UV exposure-induced skin photoaging in mice. Chin Med. 2025;20(1):146.[DOI]
-
63. Kim Y, Yoon S, Kim S, Kim Y, Jeong S, Kim HJ. Clinical efficacy of adiponectin-stimulating peptide on UV-induced skin damage. Cosmetics. 2025;12(2):54.[DOI]
-
66. Sumali SM. A comparative study of UVA and UVB radiation: Mechanisms of DNA damage and repair. Indonesia J Biomed Sci. 2023;17(2):239-243.[DOI]
-
69. Fontana GA, Singh NK, Rotankova N, Eichelberg A, Di Filippo M, MacArthur MR, et al. UVA irradiation promotes ROS-mediated formation of the common deletion in mitochondrial DNA.Life. 2026;16(4):577.[DOI]
-
73. Jomova K, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, Valko M. Several lines of antioxidant defense against oxidative stress: Antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol. 2024;98(5):1323-1367.[DOI]
-
81. Park MJ, Song NE, Song HA, Chung KS, An HJ, Hong HD, et al. Standardized hot water extract from the aerial parts of Cirsium setidens (Dunn) Nakai alleviates UVB-induced skin photoaging damage through the inhibition of MMP-1/3 and MAPK/AP-1 pathways in vitro and in vivo. J Funct Foods. 2025;129:106890.[DOI]
-
94. Notarte KIR, Quimque MTJ, Macaranas IT, Khan A, Pastrana AM, Villaflores OB, et al. Attenuation of lipopolysaccharide-induced inflammatory responses through inhibition of the NF-κB pathway and the increased NRF2 level by a flavonol-enriched n-butanol fraction from Uvaria alba. ACS Omega. 2023;8(6):5377-5392.
-
109. Wang J, Yuan M, Li Q, Shen C, Zhang X, Zhu C, et al. Combined protection against UVB-induced photoaging by oleuropein, hydroxytyrosol, and verbascoside through modulation of inflammation, oxidative stress, and collagen homeostasis. Sci Rep. 2025;15(1):41008.[DOI]
-
118. Portillo-Esnaola M, Rodríguez-Luna A, Nicolás-Morala J, Gallego-Rentero M, Villalba M, Juarranz Á, et al. Formation of cyclobutane pyrimidine dimers after UVA exposure (dark-CPDs) is inhibited by an hydrophilic extract of Polypodium leucotomos. Antioxidants. 2021;10(12):1961.
-
119. Portillo M, Mataix M, Alonso-Juarranz M, Lorrio S, Villalba M, Rodríguez-Luna A, et al. The aqueous extract of Polypodium leucotomos (fernblock®) regulates opsin 3 and prevents photooxidation of melanin precursors on skin cells exposed to blue light emitted from digital devices. Antioxidants. 2021;10(3):400.
-
121. Faisal A, Philipsen PA, Lerche CM, Douki T, Laursen FSW, Granborg JR, et al. Changes in ultraviolet B radiation-induced DNA damage and erythema after oral nicotinamide and polypodium leucotomos in healthy volunteers: An intraindividual controlled trial. Photochem Photobiol Sci. 2025;24(11):1951-1958.[DOI]
-
126. Mann T, Eggers K, Rippke F, Tesch M, Buerger A, Darvin ME, et al. High-energy visible light at ambient doses and intensities induces oxidative stress of skin-Protective effects of the antioxidant and Nrf2 inducer Licochalcone A in vitro and in vivo. Photodermatol Photoimmunol Photomed. 2020;36(2):135-144.
-
133. Threskeia A, Sandhika W, Rahayu RP. Effect of turmeric (Curcuma longa) extract administration on tumor necrosis factor-alpha and type 1 collagen expression in UVB-light radiated BALB/c mice. J App Pharm Sc. 2023;13(5):121-125.[DOI]
-
136. Ma Y, Li Y, Yao Y, Huang T, Lan C, Li L. Mechanistic studies on protective effects of total flavonoids from Ilex latifolia Thunb. On UVB-radiated human keratinocyte cell line (HaCaT cells) based on network pharmacology and molecular docking technique. Photochem Photobiol. 2025;101(1):70-82.
-
138. Shin MH, Kwon Y, Lee S, Kim HB, Kim H, Jeong S, et al. Protective effect of a novel metal-phenolic network composite against ultraviolet-induced skin damage by modulating MAPK/AP-1/NF-κB signaling pathways and attenuating oxidative stress in human keratinocytes. Biomed Pharmacother. 2025;193:118874.
-
139. Vásquez O, Contreras-Trigo B, Castillo E, Contreras N, Lemus J, Zuniga FA, et al. Chitosan nanoformulations of mycosporine-like amino acid (MAA)-rich extracts from Mazzaella laminarioides effectively protect human keratinocytes against UVA radiation damage. Int J Mol Sci. 2025;26(21):10394.
-
148. Lee E, Ahn DK, Kim JH, Lee S, Kim HJ, Lee HK, et al. Skin anti-aging and moisturizing effects of low-molecular-weight collagen peptide supplementation in healthy adults: A randomized, double-blind, placebo-controlled clinical trial. J Microbiol Biotechnol. 2025;35:e2507008.
-
159. Duteil L, Queille-Roussel C, Aladren S, Bustos X, Trullas C, Granger C, et al. Prevention of polymorphic light eruption afforded by a very high broad-spectrum protection sunscreen containing ectoin. Dermatol Ther. 2022;12(7):1603-1613.[DOI]
-
171. Vega J, Schneider G, Moreira BR, Herrera C, Bonomi-Barufi J, Figueroa FL. Mycosporine-like amino acids from red macroalgae: UV-photoprotectors with potential cosmeceutical applications. Appl Sci. 2021;11(11):5112.[DOI]
-
172. Moreira BR, Vega J, Sisa ADA, Bernal JSB, Abdala-Díaz RT, Maraschin M, et al. Antioxidant and anti-photoaging properties of red marine macroalgae: Screening of bioactive molecules for cosmeceutical applications. Algal Res. 2022;68:102893.[DOI]
-
173. Yang Y. Multi-target mechanisms and translational challenges of traditional Chinese medicine active components in anti-skin photoaging. Theor Nat Sci. 2025;129(1):134-140.[DOI]
-
185. Zhu F, Qu L, Xu R, Yuan Y, Zhang S, Chen Y. Synergistic anti-photoaging and anti-inflammatory effects of Eucommia ulmoides, Styphnolobium japonicum, and Portulaca oleracea extracts via TGF-β/Smad/IL-17 pathway. Chem Biol Technolog Agric. 2025;12(1):139.[DOI]
Copyright
© The Author(s) 2026. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
Publisher’s Note
Science Exploration remains a neutral stance on jurisdictional claims in published
maps
and institutional affiliations. The views expressed in this article are solely those
of
the author(s) and do not reflect the opinions of the Editors or the publisher.
Share And Cite



Science Exploration Style
Lai J, Ren X, Liang G, Jia S, Li J, Wang Y, et al. Research progress on skin photoaging mechanisms and natural extracts. Ageing Cancer Res Treat. 2026;3:202605. https://doi.org/10.70401/acrt.2026.0027
Tips
Copy completed.
Submit a Manuscript
Author Instructions
Cite this Article
Article Metrics
0
View
0
Download
Cited
Article Updates

- Abstract
- Keywords
- 1. Introduction
- 2. Skin Structure and Layer-Specific Responses to Photoaging
- 3. Molecular Mechanisms of Skin Photoaging
- 4. Natural Extract-Based Interventions Against Skin Photoaging
- 5. Conclusion
- 6. Challenges and Future Perspectives
- Acknowledgement
- Authors contribution
- Conflicts of interest
- Ethical approval
- Consent to participate
- Consent for publication
- Availability of data and materials
- Funding
- References
- Copyright
Science Exploration Style
Lai J, Ren X, Liang G, Jia S, Li J, Wang Y, et al. Research progress on skin photoaging mechanisms and natural extracts. Ageing Cancer Res Treat. 2026;3:202605. https://doi.org/10.70401/acrt.2026.0027
copy
Share Link
copy