Monocyte subsets across veterinary species
Javier Domínguez
1,*

,
Loems Ziegler-Heitbrock
2,*
*Correspondence to:
Javier Domínguez, Departamento de Biotecnología. Centro Nacional Instituto de Investigación y Tecnología Agraria y Alimentaria (INIA-CSIC), Madrid 2804, Spain.
E-mail: jdjuncal57@gmail.com
Loems Ziegler-Heitbrock, Independent researcher, Monocytomics Research, Herrsching 82211, Germany. E-mail: LZH@monocyte.eu
Loems Ziegler-Heitbrock, Independent researcher, Monocytomics Research, Herrsching 82211, Germany. E-mail: LZH@monocyte.eu
Myeloid Cells. 2026;1:202601. 10.70401/mc.2026.0004
Received: March 12, 2026Accepted: May 18, 2026Published: May 18, 2026
Abstract
Monocytes consist of several subsets, which differ in their phenotype and functional capacity. This heterogeneity was first shown in man, and later evidenced in other species. Here, we review the current knowledge on the phenotype and functionality of monocyte populations in species of veterinary interest, in comparison to man. The increasing availability of reagents for leukocyte phenotypic analyses, along with the growing application of -omic technologies, are enabling a better characterization of these subsets, and facilitating comparisons across species. The review demonstrates that classical, intermediate, and non-classical monocytes can be defined in pig, cattle, sheep, camel, buffalo, horse, and dog. However, more phenotypic, functional and transcriptomic studies are needed in some species to establish an accurate correspondence. A better understanding of shared and species-specific features of these monocyte subsets will pave the way for translational research on strategies selectively targeting these cells to treat different pathological conditions, including infectious, inflammatory and malignant diseases.
Keywords
Monocyte subsets, veterinary species, phenotype, surface receptors, transcriptomics
1. Introduction
Monocytes are mononuclear phagocytes, which circulate in blood and play critical roles in host defense and the maintenance of tissue homeostasis. They have long been seen to merely be precursors for macrophages; however, more recent research points to monocytes performing important functions of their own[1,2]. They act as immune sentinels, sensing danger signals, triggering inflammatory responses, and killing a broad variety of microbial pathogens, and contribute to the development and shaping of adaptive immune responses through cytokine production[3]. They also contribute to resolving inflammation and repair tissue after injury, being involved in angiogenesis, scavenging of apoptotic cells and toxic debris[4,5].
When monocytes go into various tissues, then they develop into specialized types of macrophages. During ontogeny, tissue is seeded initially by macrophages derived from embryonic precursors[6,7]. Thereafter, in homeostasis and in response to environmental and inflammatory cues, monocytes are recruited to sites of tissue remodeling and injury and differentiate into different types of macrophages. In the lung, they become alveolar macrophages[8], in the liver, Kupffer cells[9] and in the bone, osteoclasts[10]. Also, in the brain, microglia cells, which were initially proposed to only replenish by local proliferation, have been shown to be replaced by monocyte-derived cells[11,12]. In addition, this development of monocytes into macrophages is shaped by conditions like trauma, infection and malignancy[6,13].
Monocytes comprise several subsets, which differ in their phenotypes and functional capacities. The existence of monocyte heterogeneity was first evidenced in man, where two monocyte populations were initially identified based on the expression of CD14 and CD16: a major population with high levels of CD14 but no CD16 expression (CD14++ CD16-) named as “classical” monocytes (cM), which accounts for greater than 80% of total monocyte count in healthy individuals; and a minor population, which expressed the CD16 antigen on the cell surface and lower levels of CD14[14]. These CD16-positive monocytes have been further subdivided into CD14++ CD16+ intermediate monocytes (intM), which express relatively high levels of CD14 and low CD16, and CD14+ CD16++ non-classical monocytes (ncM), which express lower levels of CD14 with higher expression of CD16[15].
In mice, three monocyte subsets, can be distinguished based on the expression of Ly6C and CD43 or TREML4: Ly6C++ CD43+ TREML4- (cM), Ly6C++ CD43++ TREML4+ (intM), and Ly6C+ CD43++ TREML4++ (ncM)[15-18].
In both species, cM express high levels of CCR2 and are CD62L+, whereas ncM express low levels of CCR2 and are CD62L-[16,17,19-22]. In man, ncM also express higher levels of surface CX3CR1 than cM, as assessed by staining with a CX3CL1-Fc fusion protein[17], and confirmed by staining with monoclonal antibodies. In the mouse, a higher signal in ncM was noted in a heterozygous CX3CR1GFP/+ model[19,23], while no such difference was found for the cell surface receptor expression[24,25].
Different studies in man and mice, using various approaches such as monitoring blood monocyte repopulation after their depletion with clodronate-loaded liposomes in experimental animals[16] or with high dose chemotherapy and stem cell transplantation in man[26,27], in vivo labeling with deuterium-labeled glucose[28,29], adoptive transfer and tracking of monocyte precursors or of classical monocytes[30], or fate mapping of human classical monocytes grafted into humanized mice[28], support a developmental relationship between these subsets, with cM differentiating into intM and these subsequently maturing into ncM.
In mice, this conversion of cM into ncM appears to be driven by the interaction of Notch2 with its ligand delta-like ligand 1 (DLL1), which is expressed by a population of endothelial cells located in a specific vascular niche in bone marrow and spleen[31]. It can also be accomplished by triggering of Toll-like receptor 7 (TLR7) and nucleotide-binding oligomerization domain-containing protein 2 (NOD2) receptors with resiquimod or imiquimod, and muramyl dipeptide, respectively[32,33].
Different monocyte subsets have also been reported in several veterinary species (Table 1). However, the comparison of monocyte subsets across species with those defined in man and mouse (given at the bottom of Table 1) is hampered a) by different patterns of expression of markers like CD16, CD163 and major histocompatibility complex class II molecules (MHC II) and b) by the limited availability of reagents for phenotyping of leukocyte subsets in the different veterinary species (a selection of monoclonal antibodies (mAbs) frequently used for characterization of monocyte subsets in different veterinary species is shown in Table S1). Nevertheless, the increasing application over the last years of -omic technologies, such as bulk- and single cell transcriptomic analyses, in veterinary immunology is providing further insights in the monocyte heterogeneity of these species and has facilitated interspecies comparisons[34-36].
Table 1. Monocyte subsets in veterinary species.
| Species | Classical (cM) | Intermediate (intM) | Non-classical (ncM) | References |
| Pig | CD14++ CD163- MHC II- | CD14++/+ CD163+ MHC II- | CD14+ CD163+ MHC II+ | [41-43,45,50] |
| CD14++/+ CD163- MHC II+ | ||||
| Cattle | CD14++ CD16- CD172a++ | CD14++ CD16+ CD172a+++ | CD14- CD16++ CD172a+++ | [34,63,76,77] |
| Sheep | CD14++ CD16- | CD14++ CD16+ | CD14+/- CD16++ | [91,92] |
| Goat§ | [81] | |||
| Camel | CD14++ CD163++ MHC II+ | CD14++ CD163++ MHC II++ | CD14+/- CD163+/- MHC II++ | [96,97] |
| Buffalo | CD14++ CD163++ CD16+/- | CD14+ CD163+ CD16++ | CD14- CD163- CD16++ | [101,102] |
| Horse | CD14++ CD16- | CD14++ CD16+ | CD14+ CD16++ | [107,108] |
| Dog | CD14+/++ MHC II++ CD4- | CD14++ MHC II+ CD4+ | CD14+ MHC II- CD4+ | [110] |
| Chicken | CD115+ TIM4- | CD115+ TIM4+ | [117] | |
| Man | CD14++ CD16- | CD14++ CD16+ | CD14+ CD16++ (includes Slan+ subset) | [14,15,61] |
| Mouse | Ly6C++ CD43+ TREML4- | Ly6C++ CD43++ TREML4+ | Ly6C+ CD43++ TREML4++ | [16-18] |
§: no monocyte subsets have been defined to date; cM: classical monocytes; intM: intermediate monocytes; ncM: non-classical monocytes.
In this review, we summarize published findings in recent years on the phenotype and functionality of monocyte populations in several veterinary species (pig, cattle, sheep, goat, camel, buffalo, horse, dog, and chicken), and this is done in comparison to monocyte subsets described in man and mouse. In addition, the impact of different physiological (age) and pathological conditions (e.g., infection, stress) on the composition of these subsets is summarized. The data show that the similarity is strongest for pig, cattle and sheep, that there is good concordance for camel and buffalo and that there are many gaps for goat, horse, dog and chicken.
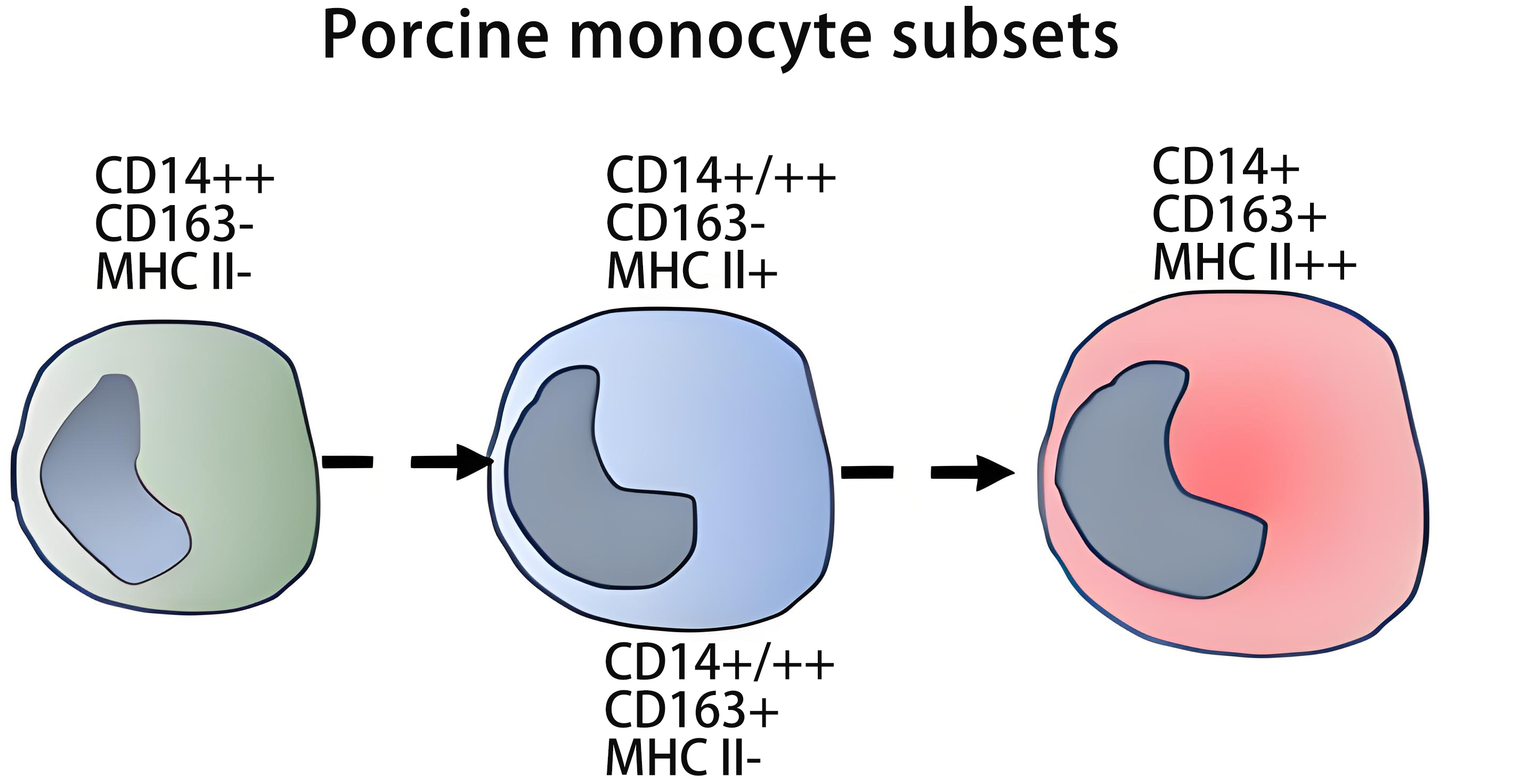
2. Monocyte subsets in the pig
Pig (Sus scrofa) monocytes can be identified among blood mononuclear cells by their high expression of CD172a[37]. Monoclonal antibodies have also been raised against porcine CD115 (CSF1R) and ADGRE1 (the homologue of mouse F4/80) that may be used as an additional monocyte marker in this species[38,39]. For a summary of porcine markers and antibodies see Table S1.
Early studies with a cross-reactive anti-human CD14 antibody showed a heterogeneous expression of this receptor in pig monocytes, with a high and low CD14 population[40]. A clear demonstration of monocyte subsets was provided by Sanchez et al.[41], and later confirmed by Ondrackova et al. and Fairbairn et al.[42,43], who described two major populations based on the expression of CD14 and CD163: CD14++ CD163- and CD14+ CD163+. These populations show approximately equal abundance in the steady-state[42,43]. These cells can be further subdivided according to the expression of MHC class II molecules into MHC II+ and MHC II- subsets. The majority of CD163- monocytes is MHC II- and almost all CD163+ monocytes are MHC II+[44-46]. However, in these studies, also CD163- MHC II+ cells and a few CD163+ MHC II- cells were described.
Regarding the CD163- MHC II+ cells, we have to consider the possibility that these might be dendritic cells. In the pig, DC1s and DC2s express high levels of CD11b[47] while monocytes are largely negative for CD11b, with only a minor subset of CD163+ cells showing low level expression of this marker[45,47]. Hence, the absence of CD11b on the CD163- MHC II+ cells suggests that they are not DCs. Furthermore, their expression of high levels of the monocyte-associated marker CD14 indicates that they are monocytes[45]. The monocyte nature of the CD163- MHC II+ cells is also supported by the absence of the DC marker CLEC12A from all monocytes, including this subset[48].
Ondrackova et al. noted CD14++ CD163- MHC II- cells (major) and CD14++ CD163- MHC II+ (minor) and CD14+ CD163+ MHC II- (minor) and CD14+ CD163+ MHC II+ cells (major)[49]. Blanc et al. in young Melanoblastoma-bearing Libechov minipigs also noted CD14++ CD163- MHC II- cells (about 30%) and CD14+ CD163- MHC II+ (about 25%) and CD14++ CD163+ MHC II- (about 6%) and CD14+ CD163+ MHC II+ cells (about 29%)[50].
This indicates that there may be 4 subsets of pig monocytes, i.e., CD163- MHC II-, CD163- MHC II+, CD163+ MHC II- and CD163+ MHC II+. Here the CD163- MHC II+ and the CD163+ MHC II- cells may represent two different types of intM (Table 1 and Table 2). Heterogeneity within intM has been noted before in man[51.52] and mouse[53].
Table 2. Phenotypic and functional characteristics of porcine monocyte subsets.
| cM | intM | ncM | References | |
| CD14++ CD163- MHC II- | CD14+/++ CD163- MHC-II+ | CD14+ CD163+ MHC-II++ | ||
| CD14+/++ CD163+ MHC-II- | ||||
| Additional surface markers | CD1a- CD11a++ CD11b- CD16+ CD49d+ CD80/86- CD172a+++ CD200R- CCR2+ (1) | CD1a- CD11a++/+++ CD11b- CD16+ CD49d+/++ CD80/86+/- CD172a+++ CD200R+/- CCR2+ | CD1a+ (subset) CD11a+++ CD11b+ (subset) CD16++ CD49d++ CD80/86+ CD172a+++ CD200R+ CCR2- | [41,43,45,46,64] |
| Transcripts (highly expressed) | CCR2, IL-10, CLEC4D, CD62L/SELL, S100As, FCN1, TREM1, LYZ | SLA DR/DQ, CD74, CD163 | CX3CR1, FCGR3A, SLA-DMA/B, ITGA4, RAP1B | [41,43,64,69] |
| FSC (size) | ++ | +++ | [41,44] | |
| Functions | IL-10 production | High T cell activation capacity, CD14+/++ CD163+ MHC-II-decrease with age | High TNF in response to LPS + IFNγ, high T cell activation capacity | [41,45,50] |
(1) Assessed by binding of rpCCL2-GFP to monocyte subsets, as well as by migration toward CCL2[64]; cM: classical monocytes; intM: intermediate monocytes; ncM: non-classical monocytes; TNF: tumour necrosis factor; LPS: lipopolysaccharide; IFNγ: interferon gamma; FSC: forward scatter.
The discrepancies regarding reports on CD163- MHC II+ and/or CD163+ MHC II- may be explained by differences in breeds and age of pigs, as well as in mAbs and gating strategies used, all which hamper comparison between studies. Differences in the levels of expression of CD14 and CD163 among breeds have been described, with monocyte subsets more clearly distinguishable in Large White x Landrace cross than in Pietrain[43]. Also, some cross-reactive anti-human CD14 mAbs (e.g., TUK4) stain lower numbers of porcine monocytes or with lower intensity compared to the anti-porcine CD14 mAb MIL2[54,55]. Likewise, when staining porcine monocytes then the use of anti-MHC II DQ mAbs results in lower number of stained cells compared to anti-MHC II DR mAbs[44,56].
In adult pigs, CD14++ CD163- MHC II- and CD14+ CD163+ MHC II+ cells represent the major subsets. These subsets correspond to the extremes of a continuum, with minor subsets in between, which exhibit intermediate expression for several markers (e.g., CD11a, CD29, CD49, CD61, CD80/86, CD200R, etc.), and which may account for the intM in this species[45,46].
As mentioned above, there may be two types of intM (CD163- MHC II+, CD163+ MHC II-) and their relative abundance may depend on age and breed. With respect to these two types of intM, we hypothesize that signals like macrophage colony stimulating factor (M-CSF), also known as colony stimulating factor 1 (CSF1) and interferon (IFN)-γ may determine which type of intM is induced. M-CSF has been shown to drive maturation of monocytes. It can induce intM in man[57] and blockade of the M-CSF pathway will reduce the number of intM[58]. Hence, this cytokine may be responsible for maturation of cM and the induction of porcine CD163+ MHC II- monocytes. On the other hand, IFN-γ can induce the co-activator CIITA, which in turn upregulates MHC class II[59]. So, this cytokine may be able to upregulate MHC class II on cM and to induce porcine CD163- MHC II+ monocytes. However, this is a hypothesis and experimental evidence is required to support this scenario.
Regarding the fate of these two types of cells, a side-by-side analysis of them has not been done. However, we suggest that both types of intM go on to become ncM. The pathways of monocyte differentiation in the pig clearly need further study.
In man, the ncM can be further defined by an antibody against the 6-sulfo-LacNAc (slan) sugar structure[60,61]. To date, no anti-slan mAb, which cross-reacts with porcine cells, has been reported, nor has any marker been identified that allows an unambiguous distinction of intermediate and non-classical populations within the continuum of staining.
Of note, CD163 shows a reverse expression pattern in porcine monocyte subsets compared to man and cattle, in that in the pig the cM are CD163-negative, while the ncM are CD163-positive[41,44,49,62]. Also, in contrast to what is seen in man and cattle, all porcine monocytes express CD16, while the more mature CD163+ cells tend to express higher levels of this molecule[41,43]. Moreover, whereas human and bovine monocytes decrease in size as they differentiate[14,63], porcine CD14+ CD163+ MHC II+ monocytes are larger than CD14++ CD163- MHC II- monocytes[41,44]. Also, the absence of MHC II from part of the porcine monocytes is clearly different from man, where all monocytes are MHC II positive.
Regarding the expression of chemokine receptors in these subsets, CCR2 and CXCR4 mRNA were clearly detected in CD14++ CD163- MHC II- monocytes but not detected or detected at low levels in CD163+ MHC II+ monocytes, whereas CX3CR1 mRNA expression was negative in CD14++ CD163- MHC II- monocytes but positive in CD163+ MHC II+ monocytes. Only the CD14hi classical monocytes migrated in response to CCL2[64]. Expression of high levels of CX3CR1 mRNA and low levels of CCR2 and CD62L mRNA by blood CD163+ monocytes has also been reported by Ondrackova et al.[65]. Hence, the chemokine receptor pattern of monocyte subsets in the pig by-and-large matches what is found in man.
In functional analysis, CD163+ monocytes produced higher amounts of tumour necrosis factor (TNF) but lower amounts of interleukin 10 (IL-10) compared to CD163- monocytes[41,45]. CD163+ monocytes were more efficient than CD163- monocytes at activating T cells against recall antigens (lysozyme) and allo-antigens[41,45].
Taking all these features into account, porcine CD14++ CD163- MHC II-, and CD14+ CD163+ MHC II+ monocytes appear to correspond to human classical CD14++ CD16- and non-classical CD14+ CD16++ monocytes, respectively. In line with this, treatment of pigs with M-CSF/CSF1, which drives the development of ncM in man, promotes an expansion of CD163+ monocytes[66]. However, transcriptomic analyses carried out by Fairbairn et al. on sorted CD14hi CD163lo and CD14lo CD163hi monocyte subsets found that whereas the former cells shared patterns with human CD14hi cM, the latter appeared to be equivalent to human intM rather than to ncM[43]. On the other hand, studies by the Dalod and Schwartz-Cornil teams found transcriptomic similarity of CD172ahi CD16hi CD163+ MHC II+ monocytes with mouse and human ncM[36]. Nonetheless, in a recent study, the Schwartz-Cornil team proposes to designate the CD16hi CD163+ subset as intermediate/non-classical monocytes, until more detailed studies (e.g., using single-cell analysis) will be carried out, since no human or mouse intermediate subsets had been included in their previous comparative transcriptomic analyses[67].
Single cell analysis may hold potential for refining the characterization of monocyte subsets in swine. An early single-cell RNA-sequencing (scRNAseq) analysis on porcine peripheral blood mononuclear cells (PBMC) discriminated five monocyte clusters (13, 19, 20, 25, 27)[68], although annotation was limited, with only a few informative differentially expressed genes (DEGs). Here cluster #19 showed the highest expression of CD14 and the lowest of CD163, and likely represents cM. Cluster #20 with intermediate levels of CD14 and CD163 and the highest levels of the MHC II encoding genes (SLA-DRA and SLA-DRB1) and of the M-CSFR (CSF1R) might represent the ncM. Cluster #13, 25 and 27 could represent intM, since they show a pattern similar to cluster #20, only that MHC class II and M-CSFR expression was somewhat lower[68]. A more recent scRNAseq analysis of PBMCs from newborn pigs by Jaudas et al. distinguished three monocyte clusters (cl_0, cl_1 and cl_2), which have been annotated as cM, intM and ncM, respectively[69]. Cl_0 showed enriched expression of CD14, S100 and LYZ transcripts; cl_1 expressed high levels of CD163, SLA-DR/DQ, and CSF1R transcripts; and cl_2 expressed the highest levels of FCGR3A/CD16, and SLA-DMA/B, and the lowest levels of CD14. Comparing cl_0, cl_1 and cl_2, there is a continuous decrease of CD14 and an increase for FCGR3A transcripts, supporting the annotation as cM, intM, and ncM. While these data are promising, we have to keep in mind that transcript levels and cell surface protein may be divergent. Hence, the data need to be consolidated by index sorting, and this requires a clear definition of intM as compared to the other subsets.
The effect of age and sex on composition of monocyte subsets have been investigated by Blanc et al. in Melanoblastoma-bearing Libechov minipigs (MeLiM) from 3 to 21 weeks of age[50]. Overall, these authors observed a decrease of CD163+ monocytes with age, mainly due to the CD14+ CD163+ MHC II- population, which showed a strong reduction after 3 months of age. Only slight differences were noted depending on sex, but females harbored a higher proportion of CD14+ CD163+ MHC II+ monocytes.
Changes in monocyte numbers with age have also been monitored by Pernold et al. in Gottingen minipigs over a period of 26 weeks under pathogen-reduced conditions[70]. The monocyte counts were high directly after birth, then dropped, followed by a rebound and then by a decrease until week 24. However, no analyses of monocyte subsets were carried out in this study even though a combination of CD172a and CD163 markers was used for staining of monocytes.
Changes in monocyte subsets have been analyzed following experimental infection of pigs with Actinobacillus pleuropneumoniae (APP), a bacterium that colonizes lungs and causes pleuropneumonia. A population of monocytes expressing high levels of CD163 but negative for MHC II (likely intM, see Table 2) was increased in the peripheral blood and bone marrow of APP-infected animals, which replaced the CD163+ MHC II+ subset (likely ncM) present on steady-state conditions[42,65]. These CD163+ MHC II- monocytes were the main producers of inflammatory cytokines, such as IL-1β, IL-6, IL-8 and TNF at the site of lung lesions[71].
In animals infected with Salmonella Enteritidis, an elevation of CD163+ MHC II+ ncM was observed in bone marrow and mesenteric lymph nodes[49].
An increase in the proportion of CD172a+ CD163+ monocytes has also been reported in piglets infected with the bacteria Glaesserella parasuis (formerly Haemophilus parasuis), the causative agent of Glasser’s disease[72].
The role of monocyte subsets has also been investigated in non-infectious disorders. Glorion et al. found a predominance of the CD16hi CD163+ MHC II+ cells (likely ncMs) within the monocytes recruited to the graft in a pig model of allogeneic lung transplantation[67]. On the other hand, in a model of cystic fibrosis in newborn piglets, Jaudas et al. reported an expansion of CD14+ CD163- MHC II- cM in lung inflammatory cell infiltrates[69].
In summary, CD172a+ CD14+ porcine monocytes can be sub-divided into four subsets, which are CD163- MHC II- cM, CD163- MHC II+ intM, CD163+ MHC II- intM and CD163+ MHC II+ ncM. Of note, there are differences to monocytes in man with respect to CD163 expression (high on ncM in the pig, low in man) and MHC II expression (absent in pig cM, present in all monocytes in man), but the pattern of an array of additional markers and many functions including cytokine production are similar to man and to other species. The major open questions relate to the nature and progeny of the two types of intermediate monocytes and to the definition of scRNAseq monocyte clusters with respect to cell type and state.
3. Monocyte Subsets in Cattle and Other Ruminants
Analysis of monocyte heterogeneity in cattle and other ruminant species has been approached by using a combination of cross-reactive antibodies, which recognize conserved epitopes across ruminant (bovine, ovine, and caprine) orthologues of CD14, CD16, CD163 and CD172a.
3.1 Cattle
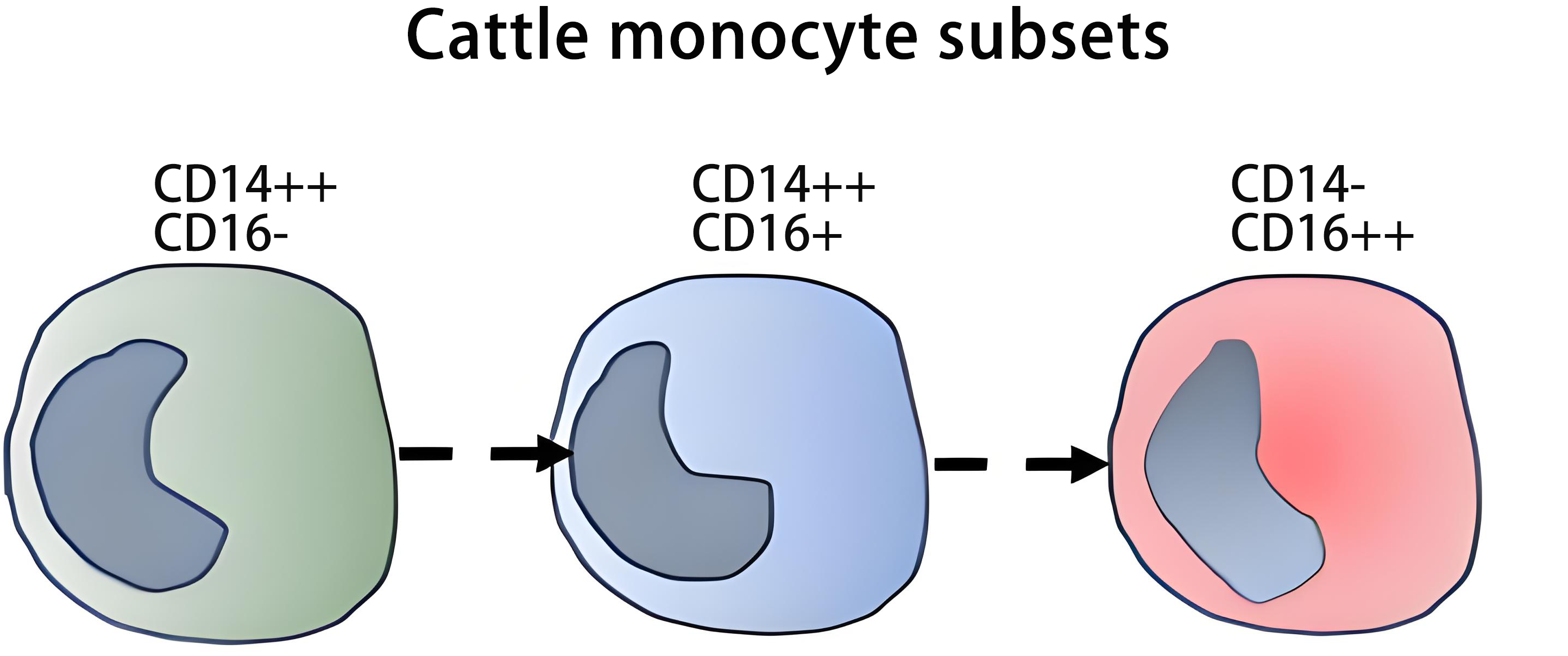
Cattle (Bos taurus) monocyte subsets have been the subject of recent reviews by Grandoni et al., and Talker et al.[73,74]. Bovine monocytes express high levels of CD172a and CD172a monoclonal antibodies like DH59B or CC149 are often used for identification of these cells in flow cytometry (Table S1).
As shown in Table 1, cattle monocytes can be divided in three subsets, phenotypically similar to those described in man, based on the expression of CD14 and CD16: CD14++ CD16- (cM), which account for about 88% of total monocytes; CD14++ CD16+, 7% (intM); and CD14- CD16++, 5% (ncM)[34,63,75-77]. The antibodies used (CAM66A and TÜK4 for CD14; KD1 for CD16) show a broad reactivity across most ruminant species (Table S1). Similar to man and mice[78], cM express the highest levels of CD62L, whereas ncM displayed the lowest CD163 expression, and intM showed the highest MHC class II expression[34,63,75]. IntM also showed the highest expression of co-stimulatory molecules CD40 and CD86, both at the mRNA and protein level[34]. Further details on patterns of cell surface markers of bovine monocyte subsets are given in Table 3.
Table 3. Phenotypic and functional characteristics of bovine monocyte subsets.
| cM | intM | ncM | References | |
| CD14++ CD16- | CD14++ CD16+ | CD14- CD16++ | ||
| Additional surface markers | CD172a ++ CD62L++ CD163+++ MHC-II ++ CD11a+ CD11b+++ CD49d+ CD40+ CD86+ CD205- CD206+/- ADGRE1+/- | CD172a +++ CD62L+ CD163++ MHC-II +++ CD11a++ CD11b++ CD49d++ CD40++ CD86++ CD205+ CD206+++ ADGRE1+ (subset) | CD172a +++ CD62L +/- CD163+/- MHC-II ++ CD11a+++ CD11b+ CD49d+++ CD40++ CD86+ CD205++ CD206++ (subset) ADGRE1++ | [34,63,74,76,77,81] |
| Transcripts | CCR2, CD163, SELL, NOD1, NLRP1, NLRC4, NLRP3, TLR4, TLR5, DEFs, S100, LYZ | MCR1, CD1E, CD74, CX3CR1, DDX58, IFIH1, BOLA-DR/DQ, CD40, CD86 | FCGR3A, MS4A7, C1QA, CX3CR1, ISGs, DDX58, IFIH1 | [34,76,77] |
| FSC (size) | +++ | +++ | ++ | [63] |
| Functions | Inflammatory responses to bacterial infection High production IL-10 High phagocytic capacity | Antigen presentation to T cells High ROS generation capacity | Antiviral responses, resolution of inflammation and tissue repair High production of TNF Low production of IL-10 | [34,63,76] |
cM: classical monocytes; intM: intermediate monocytes; ncM: non-classical monocytes; ROS: reactive oxygen species; TNF: tumour necrosis factor; FSC: forward scatter.
Functional analyses showed many similarities of bovine monocyte subsets with their human counterparts, but some differences were also noted in stimuli used and in time points. Corripio-Miyar et al. found that ncM (CD14- CD16++ cells) produced the highest levels of IL-1β, IL-12 and TNF and the lowest of IL-10 after 18h LPS stimulation, matching the patterns described in man[79], although considerable variation in cytokine levels was observed between individual animals[76]. High TNF production in ncM was also noted by Hussen et al., when looking at the mRNA level[63]. In contrast, in the same study, Hussen et al. found the lowest production of IL-1β, upon inflammasome activation after a short term (6 h) stimulation with LPS and ATP, in ncM[63]. In this study, intM were the main producers of IL-1β, as well as of reactive oxygen species (ROS) in response to opsonized bacteria or PMA, whereas cM showed the highest phagocytic capacity[63].
Most of these results have been corroborated by Talker et al. who carried out an extensive phenotypic and transcriptomic analysis of these subsets[34,77]. In line with data from Hussen et al.[63], these authors also found higher levels of IL-1β and IL-8 transcripts in cM; however, TNF transcripts were found to be primarily expressed in ncM and intM. The lowest levels of IL-10 transcripts were also found in ncM[34]. The different patterns reported in the above studies may be explained, at least in part, by differences in cell preparation, in stimuli used and in time points. Still, all of this is in line with the concept that CD16+ monocytes are superior producers of pro-inflammatory mediators, while they produce little anti-inflammatory molecules, such that the net result is a pro-inflammatory state.
The expression of chemokine receptors CCR2 and CX3CR1 in bovine monocyte subsets also matches the pattern seen in man. Classical monocytes expressed high mRNA levels for CCR2, whereas intM and ncM showed the highest expression of CX3CR1 transcripts and little or no transcripts for CCR2[76,77]. While CX3CR1 protein expression is detected in cM, with higher levels seen in intM and ncM, no data on cell surface expression of CCR2 are available[74]. Functional analysis revealed different responses of monocyte subsets to chemokines in bovine compared to human monocyte subsets. While in man cM migrate in response to CCL2, bovine cM fail to do this toward bovine CCL2[75,80]. The absence of response to CCL2 is still unexplained, and analyses of cell surface protein expression and regulation of CCR2 in bovine monocytes are needed. By contrast, bovine monocytes migrate in response to CCL5, likely via CCR1 and not CCR5, as the latter is not expressed at the transcript level, whereas CCR1 is expressed[77,80].
In bulk transcript analysis of unstimulated cells, bovine cM, compared to intM and ncM, showed highest transcript levels of genes encoding different TLRs (TLR2, TLR4, TLR5), and NOD-like receptors (NOD1, NLRP3), which suggests a prominent role of this population in inflammatory responses to bacterial infection[34,77]. In addition, cM is also high for TLR7 and 9, which are intracellular receptors involved in anti-viral defense, and this indicates that these monocytes are also important in viral infection. However, this conclusion is at variance with the finding in the same study, of the lowest response to TLR7 ligands Gardiquimod and Resiquimod in cM[34].
The enrichment in ncM and intM of transcripts for the viral RNA sensors RIG-1 and MDA-5, points towards a different role of these cells in anti-viral defense. Also, constitutive expression of interferon-induced genes (ISG15, RSAD2, IFIT1, IFIT2, IFI47, GBP 1,4,5,6) was found highest in bovine intM and ncM[34,77].
IntM, by their high expression, both at mRNA and protein level, of MHC-II and the co-stimulatory molecules CD40 and CD86, also appear to be well equipped for antigen presentation to T cells[34]. However, much of the impact of monocyte subsets on T cell function is mediated by cytokines and cell surface molecules, which are differentially expressed between cM, intM and ncM[34].
Data from bulk- and scRNAseq transcriptomic analyses as well as the pattern of expression of markers such as CD205, CD206 or CD209 are suggestive of further complexity, presumably reflecting the presence of various populations or different cell states within these subsets[34,76,77,81,82].
ScRNAseq analyses from two cows by Talker et al. revealed three clusters (c0, c3, c4) with expression signatures resembling cM, one cluster matching intM (c1) and one ncM (c2)[34]. The cM clusters were characterized by expression of S100 genes as well as ficolin and lysozyme. The expression of S100 genes decreased from c4 to c3 to c0. It further decreased in clusters c1 and c2, which is similar to what has been observed in scRNAseq analysis of human monocytes[61]. The intM cluster c1 was highest for CD1E, several MHC II genes and CD74, which encodes the MHC II associated invariant chain, in line with the bulk data in the same study.
The ncM cluster c2 showed high FCGR3A, MS4A7 and TCFL2 transcripts, which have been associated with the human scRNAseq ncM cluster[61]. In addition, C1QA, B and C expression was noted for cluster c2[34]. Several studies in man have shown that these events form a distinct cluster among FCGR3A expressing monocytes[61,83,84]. It can be expected that with higher resolution such a C1Q subcluster will also be detected in cattle monocytes.
Among the cM, c4 had very high levels of defensin gene expression. While this might reflect bona fide expression of these genes by monocytes, this result could also be explained by monocyte aggregate formation or phagocytosis of neutrophils.
The expression of CD16 on bovine cM was upregulated after 4h stimulation with IFN-γ[63], in line with the concept that cM give rise to intM. This raises the possibility that the intM in fresh blood samples, defined via the CD14+ CD16+ phenotype, might also contain such recently activated cM[74].
Wilson et al. carried out scRNAseq of PBMCs from Angus cattle and found six cM clusters, one intM cluster and one ncM cluster[85]. The cM sub-clusters all showed high CD14 and low CD16 expression. Among these, clusters cM0 and cM5 were characterized by high levels of IL1B and also of NLRP3. NLRP3 is an intracellular pattern recognition receptor of the NOD-like receptor family and is part of the inflammasome, which is crucial to the release of IL1B. This would suggest that clusters cM0 and cM5 might be particularly important in bovine inflammatory and infectious diseases. The ncM cluster showed low mRNA for CD14 and lysozyme and high mRNA for CD16 and the mannose-receptor (MR) family gene CD205 (LY75). The intM cluster showed a CD14-CD16-expression pattern in between cM and ncM and showed highest MHC II. When looking at the effects of vaccination with a commercial clostridial and leptospiral combination vaccine (Ultravac® 7in1, Zoetis), then Wilson et al. noted for the vaccine high responders a two-fold enrichment of the CD16+ monocyte cluster events.
Han et al. in a scRNAseq and single nucleus RNA sequencing (snRNAseq) transcriptome study of various tissues from 15 Holstein cows described the three monocyte clusters in blood. The cM cluster was characterized by VCAN, FCN1, LYZ and high CD14, cEBP/d and S100A genes, the ncM cluster by FCGR3A, MS4A7 and SMAD3, while the intM cluster showed intermediate levels of these DEGs[86].
Changes in monocyte subset composition have been reported in dairy cows during the peripartal period, which is associated with a higher incidence of infectious diseases like mastitis or metritis, with an increase in blood cell counts of cM, intM and ncM at day 7 after parturition, and significantly higher numbers of intM in animals with a low body condition score[87]. Moreover, higher numbers of the two CD14+ monocyte (cM and intM) subsets prior to parturition were associated with a higher probability of postpartum infectious diseases, whereas an increase in CD14−/+ monocytes (ncM) was associated with a lower risk of postpartum disease[88]. A transient increase in the number of cM and intM has also been observed after an experimental infection of calves with bovine alphaherpesvirus 1 (BoAHV-1), the causative agent of Infectious Bovine Rhinotracheitis (IBR)[89]. During lethal infection with Theileria parva, cattle show a strong increase of intM and a depletion of ncM[90].
Taken together, in cattle, similar to man, the three monocyte subsets can be clearly defined by cell surface markers and in scRNAseq. This is also true for functional patterns except for the intriguing finding that cM do not migrate in response to CCL2.
Future research should resolve the question of non-response to CCL2. Also, in scRNAseq, the annotation of the monocyte clusters needs to be standardized.
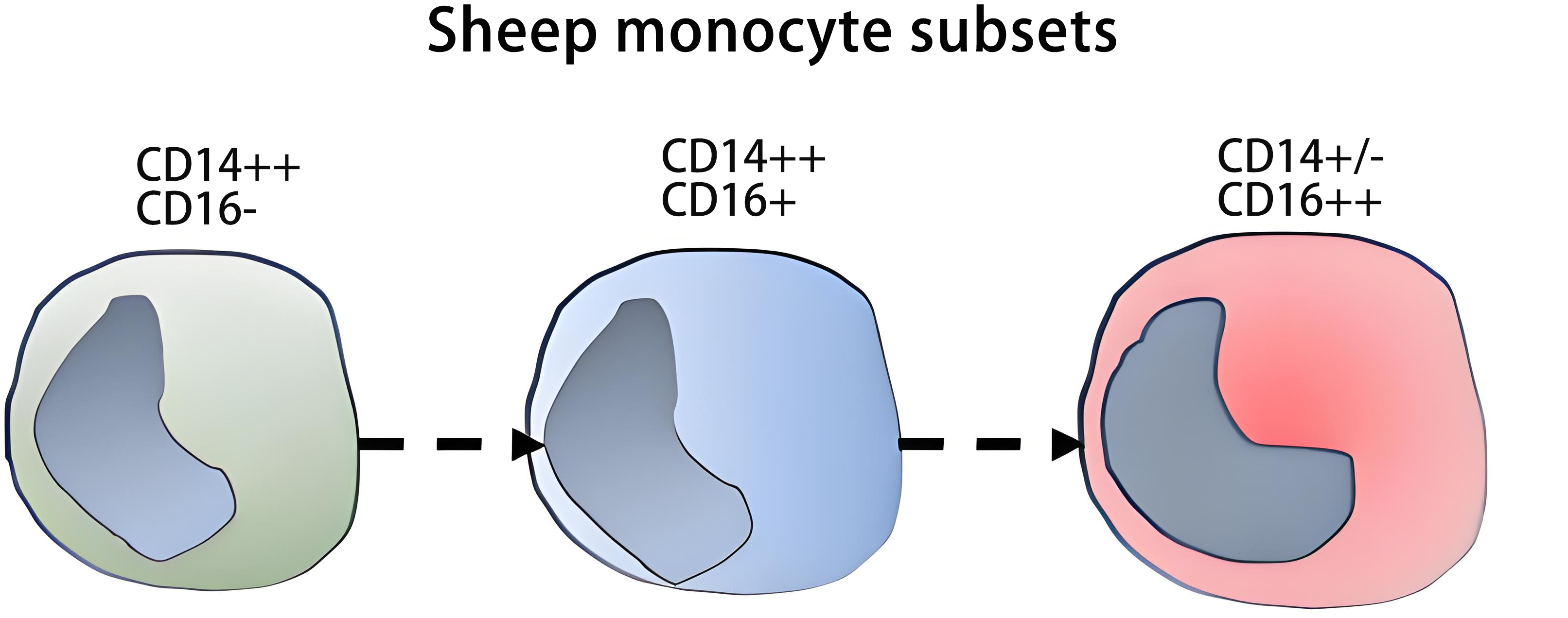
3.2 Sheep
Only a limited number of monoclonal antibodies to cell surface markers is available for sheep (Table S1). Ovine (Ovis aries) monocytes express CD172a, and according to their CD14 and CD16 expression they can be divided into three populations, CD14++ CD16-, CD14++ CD16+, and CD14+/- CD16++ (Table 1 and Table 4)[91-93], akin to cM, intM, and ncM in man and cattle.
Table 4. Phenotypic and functional characteristics of ovine monocyte subsets.
| cM | intM | ncM | References | |
| CD14++ CD16- | CD14++ CD16+ | CD14+/- CD16++ | ||
| Additional surface markers | MHC-II ++ | (no data on MHC-lI) | MHC-II +++ | [91] |
| Expression of CSF1R EGFP transgene§ | low | high | high | [91] |
| FSC (size) | ++ | ++ | ++ and +++ (two subsets) | [91] |
| Functions | Increased after vaccination against M avium paratuberculosis | [92] |
§: CSF1R-EGFP: EGFP expressed under the control of the murine CSF1R promotor plus an intronic element; cM: classical monocytes; intM: intermediate monocytes; ncM: non-classical monocytes; FSC: forward scatter; EGFP: enhanced green fluorescent protein.
In Csf1r-EGFP transgenic sheep monocytes express the reporter protein EGFP (Enhanced Green Fluorescent Protein) under the control of Csf1r promoter. In this model, ncM (CD14+ CD16++) expressed higher levels of EGFP than cM and intM (CD14++ CD16-/+ cells), which is consistent with the higher expression of Csf1r transcripts by human CD16+ monocytes compared to CD16- monocytes[91,94]. Also, in common with man, ovine CD16+ monocytes expressed higher levels of MHC II than CD14++ monocytes[91].
Ahmed et al. investigated changes in blood monocyte subsets in ewes following vaccination against Mycobacterium avium paratuberculosis[92]. Total numbers of blood monocytes and cM dropped post vaccination, whereas the numbers of ncM showed a significant increase. This is in contrast to findings in man, where vaccination against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has been shown to result in an increase of intermediate and classical monocytes[95].
Taken together, only a few studies on phenotype and function of ovine monocyte subsets have been published. Given that adequate monoclonal antibodies are available, there is great potential in the analysis of monocyte subsets in this species. Also, scRNAseq studies have not been published to date.
3.3 Goat
In contrast to sheep and other ruminants, CD14, CD16, and CD163 (Table S1) are expressed at high levels on all goat (Capra hircus) monocytes, such that no subsets can be defined with these markers[81]. There was a subset of CD14-/+ CD16+ CD163- cells but these were categorized as granulocytes based on their forward scatter (FSC) and side scatter (SSC) characteristics. Taken together, monocyte subsets in goats have not been identified to date, and studies to detect these might be successful with a broader set of antibodies or with a single cell sequencing approach.
3.4 Camel
Monocyte heterogeneity in dromedary camel (Camelus dromedarius) has been extensively investigated by Hussen and colleagues. According to the expression profiles of CD14, CD163 and MHC II, camel CD172a+ monocytes can be divided into three subsets: CD14++ CD163++ MHC II+ (≈ 87% of total monocytes), CD14++ CD163++ MHC II++ (≈ 6 % of total monocytes), and CD14+/- CD163+/- MHC II++ (≈ 5 % of total monocytes) (Table 1), which represent the counterparts of human and bovine cM, intM, and ncM, respectively[96,97]. CD14++ CD163++ MHC II++ intM display the highest phagocytic and ROS production capacity in response to bacterial stimulation[97]. These intM have been shown to expand in animals with bacterial endometritis caused by E. coli or S. aureus, and their numbers correlate with severity of the disease[98].
The composition of camel monocyte subsets has been shown to vary with age, and can also be modified by stress or infection. Newborn animals show lower numbers of CD14++ CD163++ MHC II++ intM compared to adult camels[97]. Of note, stressed camels and infected camels, with trypanosomiasis or bacterial metritis, exhibit elevated serum cortisol levels, which has been associated with a reduced CD172a, CD14 and CD163 expression on monocytes, and an increased production of reactive oxygen species[99,100].
Taken together, monocyte subsets are well defined in the camel, but there are limited functional analyses. Changes in infectious disease have been noted and such analyses can be expanded, as well as the response of monocyte subsets to exercise. scRNAseq data are not available at this point.
3.5 Buffalo
Like in cattle, monocyte heterogeneity in the water buffalo (Bubalus bubalis) has been analyzed using a combination of cross-reactive mAbs against CD14, CD16, CD163 and CD172a[101,102] (see Table S1). In contrast to cattle, almost all buffalo monocytes express CD16. Nonetheless, the pattern of expression of CD14 or CD163 vs CD16 reminds of that observed in man and cattle, with a dominant CD14++ CD163++ CD16+/- subset and two minor subsets, which are CD14+ CD163+ CD16++, and CD14- CD163- CD16++. These may represent cM, intM, and ncM, respectively (Table 1).
Grandoni et al. evaluated changes in monocyte subsets in response to an experimental infection with Bubaline alphaherpesvirus-1 in animals, which had been immunized with a live-attenuated gE-/tK-deleted marker vaccine or an inactivated vaccine[103]. An early increase in absolute numbers of cM (at days 4-7 post-challenge) was observed, followed by a late rise in intM numbers (days 15-63), which was more prominent in animals vaccinated with the live-attenuated vaccine. A significant increase in absolute numbers of ncM was also observed in animals which received the inactivated vaccine at post-challenge days 4 and 63[103]. These findings are in line with several studies in man, which demonstrated an increase in intM post vaccination with, for example, SARS-CoV2 mRNA vaccine[104] or Influenza vaccine[105].
Scata et al. investigated possible changes with age in monocyte subset composition. No differences were observed between the different age groups, (80-100 day-old buffalo calves, 16-18 month-old buffalo heifers and 4-6 year-old buffalo cows), but there were significant differences when comparing animals of same age groups from different farms[106].
Taken together, monocyte subsets in the buffalo have been defined but there are limited phenotypic and functional analysis, scRNAseq data are not available.
4. Monocyte Subsets in the Horse
Equine (Equus caballus) monocytes express high levels of CD14, as assessed by a cross-reactive anti-human CD14 mAb[107]. In a preliminary analysis with an anti-equine CD16 antibody, a minor proportion of these cells (≈ 4%) were shown to express CD16[108]. These cells might represent intM and ncM (Table 1), with the pattern of staining akin to man and cattle. A clear determination of such subsets awaits further study in a large group of animals. Here antibodies to equine CD163, CD172a and MHC II (Table S1) might help in the flow-cytometry definition of monocyte subsets in the horse.
Recent scRNAseq analysis of equine PBMC identified 4 monocyte clusters[35]. There is one cluster (# 18) with characteristics of cM, based on highest expression levels of CD14, SELL, LYZ and CD44 genes and on lowest levels of MHC II, DRA and DQA. Then there are two clusters (# 11 and #12), which display intermediate levels of these genes (CD14, CD44, SELL, MHC II DRA and DQA) but the highest level of CD163 transcripts. Finally, there is cluster #28, which accounts for less than 6% of total monocytes, and expresses high levels of CD16, CX3CR1, NR4A1, HES4, and MHC II DRA and DQA, in line with the pattern of ncM in man[35].
These data suggest that the cM/intM/ncM-classification can also be applied to horse monocytes. A more comprehensive flow cytometry approach should be able to define these cell populations via cell surface markers. Also, further experiments are required to assign functions to these different subsets.
5. Monocyte Subsets in the Dog
Canine (Canis lupus familiaris) monocytes can be gated according to the expression of CD11b and the lack of the neutrophil marker CAD48A. Based on the expression of CD14 and MHC II molecules, Gibbons et al. distinguished three subsets of canine monocytes: CD14+ MHC II-, CD14+ MHC II+ and CD14- MHC II+, and surmised that, by analogy with the higher expression of MHC II molecules found in CD16+ monocytes in man, they might be the counterparts of human cM, intM, and ncM, respectively[109].
Rzepecka et al. defined dog monocytes as CD11b+ CD14+ cells and they characterized three subsets of canine monocytes, which differ in the expression of MHC II and CD4 molecules: MHC II+ CD4- (which represent about 75% of the whole population), MHC II+ CD4+ and MHC II- CD4+ (comprising ≈ 12 % each one)[110] (Table 1). No CD14 negative population was detected in this work, in contrast to the study by Gibbons et al., where the proportion of CD14-negative cells ranges from 3% to 31% of total monocytes, despite the use of the same anti-CD14 antibody (mAb Tük 4). Both CD4+ subsets produced higher basal levels of ROS than the CD4- subset. The MHC II- CD4+ cells displayed the lowest levels of CD14, and were the smallest in size (and granularity), and might correspond to ncM.
More recently, MacDonald et al. have also defined in flow cytometry three canine monocyte subsets[111]. The majority of cells showed high CD14 and CD11b and were suggested to be cM. Then, there were CD14+CD11b+ cells and finally CD4++ monocytes with lowest CD49d expression. Given the limited overlap of markers in the above studies, it is difficult to align the monocyte subsets and further comprehensive studies are awaited. Of note, the CD4++ monocytes were found to be significantly increased in chronic skin inflammation[111]. This suggests that the study of monocyte subsets in dogs may be of clinical use for diagnosis and therapy.
A single cell transcriptomic analysis of canine blood leukocytes identified 9 monocyte clusters[112]. In this study, CD4+ monocytes were regarded to be equivalent to cM, and CD4- monocytes to ncM. However, CD4 expression was based on transcript levels and the annotation was limited, such that markers employed for definition of monocyte subsets in other species were not available at this point. Updates of the canine reference genome may improve the analysis of canine monocyte subsets at the scRNAseq level. In this regard, a recent scRNAseq analysis of PBMC from 6 dogs (Maltese, Poodle, mixed breed) using a new canine genome reference (canFam4) described 2 clusters of monocytes, one with high CD14 and one with high expression of CD83[113]. The CD14+ cluster contained a CCL3 high CCL4 high sub-cluster and a CD4 expressing sub-cluster. A clear definition of cM, intM and ncM was not possible in this study.
Further transcriptomic and functional studies will help to establish accurate correspondences with the monocyte populations described in other species.
6. Monocyte Subsets in the Chicken
Chicken (Gallus gallus) blood monocytes can be identified using a mAb against CD115/CSF1R (ROS-AV170) and the mAb KUL01, which recognises MRC1L-B, a homologue of the mammalian mannose receptor C-type 1 (MRC1, CD206)[114-116]. Hu et al. studied CSF1R-mApple transgenic chicken, whose monocytes express the red fluorescent protein mApple under the control of CSF1R regulatory sequences[117]. They showed that these cells can be divided into two populations, roughly equivalent in numbers, based on the expression of TIM4, which is a phosphatidylserine receptor (Table 1). All monocytes are MHC II+, but TIM4+ cells compared to TIM4- cells show higher levels of CX3CR1 and C1QA transcripts, such that they match human ncM (Table 1). These cells also show a 1.4-fold higher expression of the MHC II associated invariant chain CD74 and higher transcripts encoding receptors involved in apoptotic cell recognition and removal such as MARCO, MERTK, LRP1 or STAB1. This led to the suggestion that these TIM4+ monocytes may act as resident scavengers of blood[117].
Also, in a recent scRNAseq analysis by Maxwell et al., monocytes were defined as mannose-receptor mRNA-positive cells and 2 clusters (CL5: MR-high MHC II-low S100A12-high, and CL6: MR-low MHC II-high S100A12-low) were described[118]. These may represent cM and intM/ncM cells, respectively. Re-clustering identified 2 sub-clusters of CL5 (#0,1) and 3 sub-clusters of CL6 (#3,4,5). Sub-cluster #1 was high for S100A12 and the serine proteinase inhibitor SPINK2, while subcluster #5 showed high transcript levels for CD81 and SAA and for MHC II. An alignment of the main monocyte clusters in Hu et al.[117] and Maxwell et al.[118] studies is difficult because of the limited overlap of informative DEGs between them. However, the higher expression of CD74 for TIM4+ cells[117] and the higher MHC II for cluster CL6[118] suggest that the TIM4+ cells and the cluster CL6 might overlap.
These scRNAseq data provide a first glimpse into the heterogeneity of chicken monocyte subsets and further studies may provide a clearer picture of cM, ncM and maybe additional clusters and populations. Another open question is whether a population of intermediate monocytes exists in the chicken.
7. Transcriptional Control
Multiple signalling pathways and transcription factors have been shown to control development and gene expression in monocytes in man and mouse, and this includes the Notch signalling pathway[31,119], c/EBPb[119] and NR4A1[120]. By contrast, to our knowledge, no specific data on transcriptional control of monocyte subset development by these or other transcription factors are available for the veterinary species covered in this review.
However, a core regulatory complex (C/EBPb, IRF8, KLF2, KLF4, and NR4A1), which has the potential to control development of ncM, is conserved across species[121]. This includes the mammalian species covered in this review, while it applies to birds to a lesser extent.
Furthermore, much of the monocyte biology is governed by M-CSF (CSF-1), and the regulation of this gene is controlled by conserved elements in the second intron of this gene. This fms intronic regulatory element, includes binding sites for transcription factors PU.1, AP1, KLF4, RUNX1 and CEBP and is conserved in pig, cattle, sheep and chicken[122]. This indicates that there may be conserved transcriptional control, but specific data for veterinary monocytes are needed.
8. Conclusions and Prospects
Monocyte subsets from veterinary species share many similarities in phenotype and function with those from man and mouse, reflecting evolutionary conservation of monocyte heterogeneity. However, important differences between species in the expression of surface receptors (e.g., CD16, CD163, MHC II) and functional capacities (response to chemokines, cytokine production after inflammasome activation, ROS generation, NO production, etc.) have also been noted, which demand caution when attempting to extrapolate observations made in one species to another[34,36,43,75,78].
Despite the remarkable progress made in the last decades in the study of the immune system of veterinary animals[123,124], the veterinary toolbox for the study of the immune system is still limited compared to what is available for man and mouse. This has led to the use of different combinations of markers for phenotypic characterization of monocyte subsets in different species. This together with interspecies variations in the pattern of expression of some markers (e.g., CD16, CD163) and the shortage of functional studies, have hampered their comparison with the subsets defined in man and mouse.
However, the increasing use of -omic techniques, and particularly of bulk- and single cell transcriptomic analyses in veterinary immunology, with a growing number of species with sequenced and annotated genomes, is helping to overcome these limitations, as they allow us to define the signatures of the different cell populations. The transcriptome analysis of single cells and of purified bulk populations can reflect the functional capacities of monocyte subsets and is contributing to a better delineation of the relationships of monocyte subsets across species. These transcriptomic analyses may also lead to the identification of novel cell surface markers that could facilitate the discrimination of new subsets or cell states, and that could be used for their isolation and deeper analysis.
Advances in the characterization of monocyte subsets in veterinary species will provide insights into the mechanisms of pathogenesis of inflammatory and infectious diseases affecting the health and productivity of these species. Some of them (such as pig, sheep, and cow) are also excellent models for translational research in a variety of human diseases, where monocytes play an important role in the pathogenesis[125-128]. For this, a better understanding of how monocyte subsets in veterinary species correlate with those of man will pave the way for the use of these animals for testing therapeutic strategies that target specific monocyte subsets.
Supplementary materials
The supplementary material for this article is available at: Supplementary materials.
Authors contribution
Domínguez J, Ziegler-Heitbrock L: Conceptualization, writing-original draft, review & editing.
Conflicts of interest
The authors declare no conflicts of interest.
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Availability of data and materials
Not applicable.
Funding
None.
Copyright
©The Author(s)2026.
References
-
14. Passlick B, Flieger D, Ziegler-Heitbrock HW. Identification and characterization of a novel monocyte subpopulation in human peripheral blood. Blood. 1989;74(7):2527-2534.[PubMed]
-
30. Hettinger J, Richards DM, Hansson J, Barra MM, Joschko AC, Krijgsveld J, et al. Origin of monocytes and macrophages in a committed progenitor. Nat Immunol. 2013;14(8):821-830.[DOI]
-
37. Alvarez B, Sánchez C, Bullido R, Marina A, Lunney J, Alonso F, et al. A porcine cell surface receptor identified by monoclonal antibodies to SWC3 is a member of the signal regulatory protein family and associates with protein-tyrosine phosphatase SHP-1. Tissue Antigens. 2000;55(4):342-351.
-
41. Sánchez C, Doménech N, Vázquez J, Alonso F, Ezquerra A, Domínguez J. The porcine 2A10 antigen is homologous to human CD163 and related to macrophage differentiation. J Immunol. 1999;162(9):5230-5237.[PubMed]
-
56. Bullido R, Doménech N, Alvarez B, Alonso F, Babín M, Ezquerra A, et al. Characterization of five monoclonal antibodies specific for swine class II major histocompatibility antigens and crossreactivity studies with leukocytes of domestic animals. Dev Comp Immunol. 1997;21(3):311-322.
-
57. Weiner LM, Li W, Holmes M, Catalano RB, Dovnarsky M, Padavic K, et al. Phase I trial of recombinant macrophage colony-stimulating factor and recombinant gamma-interferon: Toxicity, monocytosis, and clinical effects. Cancer Res. 1994;54(15):4084-4090.[PubMed]
-
58. Korkosz M, Bukowska-Strakova K, Sadis S, Grodzicki T, Siedlar M. Monoclonal antibodies against macrophage colony-stimulating factor diminish the number of circulating intermediate and nonclassical (CD14(++)CD16+/CD14+CD16(++)) monocytes in rheumatoid arthritis patient. Blood. 2012;119(22):5329-5330.
-
69. Jaudas F, Bartenschlager F, Shashikadze B, Santamaria G, Reichart D, Schnell A, et al. Perinatal dysfunction of innate immunity in cystic fibrosis. Sci Transl Med. 2025;17(782):eadk9145.[DOI]
-
85. Wilson A, Alexandre PA, Brice AM, Hine BC, Ingham A, Legrand TPRA, et al. Single-cell transcriptomics uncovers key immune drivers of vaccine efficacy in cattle. BMC Genom. 2025;26(1):750.[DOI]
-
88. Pomeroy B, Sipka A, Hussen J, Eger M, Schukken Y, Schuberth HJ. Counts of bovine monocyte subsets prior to calving are predictive for postpartum occurrence of mastitis and metritis. Vet Res. 2017;48(1):13.[DOI]
-
94. Mobley JL, Leininger M, Madore S, Baginski TJ, Renkiewicz R. Genetic evidence of a functional monocyte dichotomy. Inflammation. 2007;30(6):189-197.[DOI]
-
103. Grandoni F, Hussen J, Signorelli F, Napolitano F, Scatà MC, de Donato I, et al. Evaluation of hematological profiles and monocyte subpopulations in water buffalo calves after immunization with two different IBR marker vaccines and subsequent infection with Bubaline alphaherpesvirus-1. Vaccines. 2023;11(9):1405.
-
106. Scatà MC, De Matteis G, Grandoni F, Di Vuolo G, Cappelli G, Vecchio D. Characterization of cellular immune system at different ages in water buffalo (bubalus bubalis). J Buf Sci. 2024;13:133-139.[DOI]
-
114. Garcia-Morales C, Rothwell L, Moffat L, Garceau V, Balic A, Sang HM, et al. Production and characterisation of a monoclonal antibody that recognises the chicken CSF1 receptor and confirms that expression is restricted to macrophage-lineage cells. Dev Comp Immunol. 2014;42(2):278-285.
-
121. Araujo DJ, Fernandez D, Alimadadi A, Zagal-Norman A, Hedrick CC. Modeling the evolutionary history of nonclassical monocytes in mammals. bioRxiv [Preprint]. 2022.[DOI]
Copyright
© The Author(s) 2025. This is an Open Access article licensed under a Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, sharing, adaptation, distribution and reproduction in any medium or format, for any purpose, even commercially, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
Publisher’s Note
Science Exploration remains a neutral stance on jurisdictional claims in published
maps
and institutional affiliations. The views expressed in this article are solely those
of
the author(s) and do not reflect the opinions of the Editors or the publisher.
Share And Cite



Science Exploration Style
Domínguez J, Ziegler-Heitbrock L. Monocyte subsets across veterinary species. Myeloid Cells. 2026;1:202601. https://doi.org/10.70401/mc.2026.0004
Tips
Copy completed.
Submit a Manuscript
Author Instructions
Cite this Article
Article Metrics
0
View
0
Download
Cited
Article Updates

Science Exploration Style
Domínguez J, Ziegler-Heitbrock L. Monocyte subsets across veterinary species. Myeloid Cells. 2026;1:202601. https://doi.org/10.70401/mc.2026.0004
copy
Share Link
copy